info@biomedres.us
+1 (502) 904-2126
One Westbrook Corporate Center, Suite 300, Westchester, IL 60154, USA
Site Map
Received: November 21, 2022; Published: November 28, 2022
*Corresponding author: Nowinszky L, University Professor at Eötvös Loránd University, Savaria Campus, Savaria Science Centre, H-9701 Szombathely, Károlyi Gáspár Square 4, Hungary
DOI: 10.26717/BJSTR.2022.47.007482
Values of the solar mean magnetic field were related to catch data for 40 taxa of Lepidoptera from a light trap in Tasmania, Australia operated for decades. This supplemented recent analyses of light trap data from three continents that revealed relationships between insect activity and many factors of the Sun and Moon. Magnetic field values, with corresponding relative catch values, were grouped into several classes. Within each group three point weighted moving averages of relative catch values were calculated. Almost half of the taxa showed a positive relationship between relative catch and absolute values of the solar mean magnetic field while one quarter showed a negative relationship.
Keywords: Insect Activity; Insect Flight; Solar Activity
Abbreviation: SMMF: Solar Mean Magnetic Field as measured by the Wilcox Solar Observatory; RC: relative catch as explained in the methods section
Catch data from light traps are assumed to reflect the actual flight activity of insects to a greater or lesser degree. Analyses of Australian light trap data have mostly examined the effects of weather and climate on the abundance and migration of insects although a few studies examined lunar factors (phase and illuminated fraction). These were reviewed by Hill [1]. In recent years, Nowinszky, et al. [2,3]and Puskás, et al. [4] used data from three continents including Australia to reveal relationships of light and pheromone trap catches to many lunar and solar factors such as lunar phase, illumination lux, illuminated fraction of the Moon, gravitational pull of the Sun and Moon, position of the Arago and Babinet points, polarization by the Sun and the Moon, sunspot numbers, solar 10.7 cm radio flux index, solar Q index and solar flare activity numbers.
Dreyer, et al. [5]used wild, tethered Bogong moths Agrotis infusa in flight simulators to investigate the influence of Earth’s magnetic field and horizontal landmarks on flight direction. The steering direction of Bogong moths was concluded to be, ‘the result of an interaction between visual landmarks and the earth’s magnetic field. This species is included in the current study. The significance of migration to pest prognosis has been much studied in Australian agricultural entomology. Farrow and McDonald [6]reviewed migra tion strategies of noctuid pests in Australia including species in the noctuid genera Agrotis, Chrysodeixis, Heliothis, Helicoverpa, Mythimna, Persectania and Spodoptera. They proposed that migration enables these species to track host plants and erratic rainfall across the continent. Five such species are included in the current study. Hill (2007) noted that a large proportion of the biomass of moths light-trapped at Stony Rise in Tasmania in spring are known or suspect migrants as did Drake, et al. [7] first note for earlier Tasmanian light traps at other locations.
This proportion decreases in summer [8]. However, the residential versus migrant status of many insects trapped at Stony Rise remains unclear and probably spans a spectrum of behaviour rather than binary alternatives. This study sought to identify common responses to solar activity among closely related species of Tasmanian Lepidoptera or those with similar migratory life strategies.
Data Sources
In our study we used the data of mean magnetic field of the Sun published by the Wilcox Solar Observatory from 1975 to 2020. These solar mean magnetic field (SMMF) values are averages of the solar longitudinal component of the Sun’s photospheric magnetic field, averaged over the entire visible disk of the Sun and taking into account the instrument profile [9]. Positive and negative values indicate the direction of the mean field (+: north polarity, -: south polarity). In this report we illustrate the results of the analysis in terms of the absolute values of the SMMF. Data on catches of moths derive from a single light trap operated at Stony Rise near Devonport in Tasmania, Australia from 1992 until 2019. The trap was typically operated every night, with some exceptions as detailed by Hill [10], with all individuals of selected taxa counted. Sometimes the catch was not emptied daily so that a single ‘trapping event’ represents variously 1-14 nights of trapping. There were 5433 trapping events embracing 7897 nights including 4167 single-night events, 502 two-night events and 516 three-night events. Data from 248 events of variously 4 to14-nights duration were not used. Data from some collecting events was not used because corresponding SMMF data was unavailable.
Data Availability
The raw catch data is available in Excel spreadsheet format from the leading author’s Research Gate webpage (https://www. researchgate.net/project/Long-term-light-trap-data-for-insectsin- Tasmania-1992-2019). This dataset is also available in other formats at https://doi.org/10.5281/zenodo.6820319 and at https:// www.gbif.org/dataset/044f96bc-3bf2-4a38-9f7c-8808ab48dbf1# contacts.
Statistical Analysis
Basic data were the number of individuals of each species caught per night drawing on catches from single, double and triple night collecting events. In order to compare the differing sampling data, relative catch values were calculated. The relative catch (RC) was defined as the quotient of the number of specimens caught during a sampling time unit (one night) per the average nightly catch of individuals within a calendar year sampling period. The RC equals one when the catch for one particular night equals the average nightly catch for that year [11]. Collecting events with nil catch of a particular species were excluded from the analysis for that species. Relative catches were grouped into classes. The number of these classes was calculated using the formula of Sturges (Odor and Iglói 1987): k = 1 + 3.3* 1g n, where k = the number of classes and n = the number of observation data.
It is not reasonable to have big differences in the number of data across classes. Therefore, the classes at the two extremities are wider than those in the middle. Within each class we used our own method and calculated three point weighted moving averages from the values of the dependent variable [11]. In preceding studies, the use of moving averages meant that the first and last values, which often carry valuable information on the most important biological impacts, were lost. In elaborating our method, we considered the work of Urmantsev [12]. He came up with a solution to ensure that no data is lost, with every initial data being accompanied by a moving average value. The new method assigns differing weights to the middle, previous and following values. Thanks to this method, our moving averages get weighted with the number of initial data. The three point moving average is calculated on the basis of the following formulae:
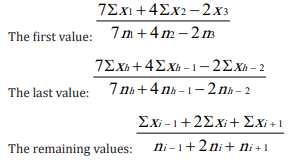
Where n = the number of catch data for the given nights and h = the designation of the additional nights. The use of moving averages is justified whenever the independent variable is made up of data representing a wide range of values that are to be contracted into classes. The dividing line between these classes is always drawn more or less arbitrarily. Extreme values in two neighbouring classes of the independent variable are always closer to each other than they are to the middle value of their own class. Working with moving averages ensures a degree of continuity between the data of our arbitrarily established classes and partly eliminates the disturbing influence of other environmental factors that are not being examined in the analysis [11]. In this work, we chose a slightly different solution. Separately, all catch data by species per year were considered as a single sample and thus relative catch values were calculated. This solution also made it possible to determine the effectiveness of trapping from the relative catch values of each year and to compare the effectiveness of the years.
The results were plotted for each of 40 taxa using both the polar and absolute values of the SMMF but individual plots are not given here. Instead responses were classified as positive, negative or ambivalent in regard to absolute values of the SMMF (Table 1) and pooled plots for taxa within each of the three types of relationship are given, with the ambivalent category divided into two subcategories (Figures 1-4). However, the RC values with associated positive and negative values of the SMMF are tabulated for all 40 taxa in Supplementary Information (Table 1). Positive correlation of RC to the absolute value of the SMMF occurred in 18 of the 40 taxa studied or 45% of taxa (Figure 1). Negative correlation of RC to absolute SMMF occurred in 10 of the 40 taxa (25%). The negative correlations were perhaps clearer than the set of positive correlations (Figure 2). The relationships were unclear or ambivalent in 12 taxa (30%), which can be split into a group of eight with an ambivalent positive relationship and a group of four with an ambivalent negative relationship (Figures 3-4).
Table 1: Relationships of RC of 40 taxa of Lepidoptera in Tasmania to the absolute values of SMMF with their taxonomic identity, number of specimens caught, number of catching nights and the year in which catching commenced. +ve means positive relationship,-ve means negative relationship.

Figure 1 Pooled chart of 18 individual taxa that showed a positive relationship of RC to the absolute value of the SMMF. One outlying coordinate (out of 133 coordinates) was excluded. This was a very high RC of 2.745 for Metasia capnochroa at the SMMF value of +33.56 micro-Tesla.

Figure 2 Pooled chart of 10 individual taxa that showed a negative relationship of RC to the absolute value of the SMMF.

Figure 3 Pooled chart of 8 individual taxa that showed no clear relationship of RC to the absolute value of the SMMF. The hollow markers are 8 outlying coordinates, one per taxon, representing low RC values at high positive (4) or negative (4) values of SMMF.

Figure 4 Pooled chart of 4 individual taxa that showed no clear relationship of RC to the absolute value of the SMMF. The hollow markers are 4 outlying coordinates, one per taxon, representing high RC values at high positive values of SMMF.

Potential relationships in the ambivalent group of 12 taxa were each confounded by a single contrary RC response at the highest value of SMMF. In the case of the group of four taxa, this value was always a positive SMMF value. In the case of the group of eight taxa, the confounding coordinate had a high positive SMMF value in four instances and was negative in four instances.
The influence of the solar magnetic field on the interplanetary magnetic field and that of the Earth is complex [13,14]. The coincidence or delay in the appearance of a terrestrial phenomenon depends on whether electromagnetic or corpuscular radiation is the cause. Such events include changes in the ionosphere and the upper atmosphere’s magnetosphere, the formation of weather fronts and sudden changes in the characteristics of ground magnetism. The basic phenomenon underlying many solar factors is the complex solar magnetic field.
Closely related taxa did not necessarily show the same relationship of RC to SMMF (Table 1), whether expressed in absolute or polar values. Nevertheless, five Noctuidae in or near the genus Proteuxoa displayed a broadly similar positive response and four of the five noctuine Noctuidae had a positive relationship of RC to absolute SMMF. The exception was the migratory Agrots munda, which shared a negative response with the migratory hadenine noctuid moth, Persectania ewingii. The three crambid species of Hednota showed three different relationships of RC to absolute SMMF. Similarly, not all migratory species shared the one and same relationship of RC to absolute SMMF. For example, positive relationships occurred in P. xylostella, A. infusa and H. punctigera, negative relationships in U. pulchelloides, C. argentifera, P. ewingii and A. munda and no clear relationship occurred in E. behrii. In regard to polar values of SMMF, these eight well understood migratory species exhibited one positive relationship (E. behrii), four increasing to decreasing relationships and three decreasing to increasing relationships.
This study found that the flight activities of 70% of the 40 moth taxa studied, as indicated by prolonged captures at one light trap, show a positive or negative relationship to the solar mean magnetic field when expressed in absolute values. Nowinszky, et al. [3] also found comparable relationships for eight species of Lepidoptera and Trichoptera in Hungary and the USA although they used polar values of SMMF. The cause of these relationships is not understood but recent studies have shown relationships of flight activity to particular solar activities such a flares, sunspots and F10.7 cm radio flux, which are known to have effects on terrestrial weather (e.g. Puskás, et al. [14].
Analysis of large sets of data from multiple traps is likely to confirm that the solar magnetic field has an impact on the activity of insects. Nowinszky, et al. [2] found in studies of various lunar and solar factors using catch data from many traps located in Hungary, Tasmania and the USA that data from a single or a few light-traps had standard deviations that were large because of the significantly different catches on different days. The standard deviations are also extremely large for migratory species that appear intermittently in highly variable numbers. The current study used a single trap whose catch included a high proportion of migrants (perhaps 40%). The use of a three point moving average helped to mitigate this problem. Long term catch data is rare but data for some pestiferous noctuid taxa in mainland Australia exists.
The authors are grateful to Dr Donald Hobern for formatting the Tasmanian light trap data for internet access through Zenodo and the Global Biodiversity Information Facility although the data used here came directly from the leading author.
(Table 1) of Supplementary Information provides the polar values of the solar mean magnetic field for all of the 40 taxa.

























