 Mini Review
Mini ReviewMini Review
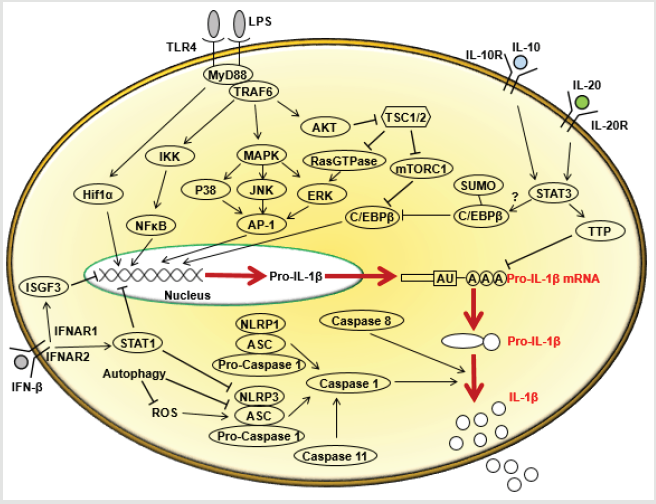
Figure 1: Multiple pathways regulating IL-1β production in macrophages.
Note: In response to LPS, pro-IL-1β transcription is activated and then is cleaved to produce the biologically active IL-1β. NF- κB and MAPK pathways are critical for the regulation of IL-1β transcription. LPS-induced phosphorylation of AKT leads to the phosphorylation and activation of CREB that binds to C/EBPβ promoter and mediates C/EBPβ expression. STAT3 pathway activated by IL-20 leads to SUMO-mediated inactivation of C/EBPβ. C/EBPβ protein is required for the activation of IL-1β gene expression. IL-10 initiates TTP expression, which could bind to IL-1β 3’UTR and facilitate its degradation. Intracellular succinate acts as an endogenous danger signal to stabilize HIF-1α, which in turn promotes IL-1β expression. The maturation of pro-IL-1β into the active form of IL-1β is thought to be mainly mediated by caspase-1, which could be activated from pro-caspase-1 by NLRP1 and NLRP3 inflammasomes. Besides, caspase-11 and caspase-8 also play important roles for the IL- 1β processing and activation. TSC1/2 is an endogenous negative regulator of mTORC1. Over-activated mTORC1 caused by TSC1/2 deficiency leads to decreased pro-IL-1β expression by inhibiting C/EBPβ expression. However, the absence of TSC1/2 increases the activity of RasGTPase, which leads to the activation of ERK and then potentially induces IL-1β transcription. Therefore, TSC1/2 controls IL-1β transcription through multiple pathways.
As an important pro-inflammatory cytokine, IL-1β exerts its
pro-inflammatory function by promoting inflammatory cytokines
production and recruiting inflammatory cells to the site of injury
or infection from the circulation.IL-1β is implicated in many
pathological conditions, including acute inflammation, chronic
inflammation and autoimmune diseases. Blocking IL-1β has been
applied to cure patient with a class of inflammatory syndromes
such as aphthous stomatitis, pharyngitis, rheumatoid arthritis,
type 2 diabetes, urticarial vasculitis and so on [1]. Therefore, it
is important to study the signaling pathways that regulate IL-1β
production. In response to an exogenous stimulus such as LPS, IL-1β
transcription is first activated. Then IL-1β is translated as an inactive
precursor protein pro-IL-1β, which need to be further cleaved at
the N-terminal 116 amino acids to produce the biologically active
IL-1β. The regulation of IL-1β production is mainly processed
on these two aspects (Figure 1). It is well known that NF-κB and
Mitogen-Activated Protein Kinase (MAPK) signaling pathways are
important for the regulation of IL-1β transcription [2,3]. LPS-TLR4
phosphorylates IKK through a Myeloid Differentiation Primary
Resposnse Gene 88 (MyD88) dependent pathway, which releases
NF-κB into the nucleus to regulate IL-1β transcription. In addition,
MyD88 signaling also participates in the activation of Transforming
growth factor-β-Activated Kinase 1 (TAK1), which results in the
activation of MAPK cascades. MAPK can active downstream signals
including P38, ERK and JNK, which activates transcription factor
activator protein 1 (AP-1). Blocking any of P38, ERK and JNK
activity could inhibit IL-1β transcription significantly [3,4].
CCAAT/enhancer-binding protein β (C/EBPβ) protein is
required for the activation of mouse and human IL-1β gene
expression [5]. C/EBPβ expression could be induced by LPS. LPSinduced
phosphorylation of AKT can lead to the phosphorylation
and activation of cAMP response element B (CREB) that binds to C/
EBPβ promoter and mediates C/EBPβ expression. STAT3 pathway
activated by IL-20 could lead to SUMO-mediated inactivation of
C/EBPβ, which interferes with the induction of IL-1β expression
in keratinocytes [6]. In macrophages, IL-10 initiates an STAT3-
dependent increase of RNA-destabilizing factor tristetraprolin
(TTP) expression. TTP could bind to IL-1β 3’UTR and facilitate its
degradation by initiating the assembly of RNA decay machinery
[7]. In addition, macrophages activated by LPS switch their core
metabolism from oxidative phosphorylation to glycolysis. Chronic
activation of macrophages by LPS causes an accumulation of
intracellular succinate by a glutamine-dependent pathway.
Succinate acts as an endogenous danger signal to stabilize hypoxiainducible
factor-1α (HIF-1α), which in turn binds to the -300
position of the IL-1β promoter and specifically promotes IL-1β
expression [8]. Besides the regulation of IL-1β expression at the
transcriptional level, protein maturation step is also important to
produce the functional IL-1β. The maturation of pro-IL-1β into the
active form of IL-1β is thought to be executed by canonicalcysteine
protease caspase-1. Therefore, the regulation of IL-1β activity is also
influenced by the regulation of caspase-1 activity. Caspase-1 could
be activated from pro-caspase-1 by NACHT, LRR and PYD domainscontaining
protein 1(NLRP1) and NLRP3 inflammasomes [9]. ASC
is one component of both NLRP1 and NLRP3 inflammasomes.
Autophagy could recognize ubiquitinated ASC and induce
selective degradation of inflammasomes and thereby suppress the
production of IL-1β [10]. NLRP3 inflammasome can be activated
by ROS stimulation. Autophagy can decrease ROS level through the
turnover of old and dysfunctional mitochondria. Accumulated old
and dysfunctional mitochondria produce an excess of ROS upon
stimulation [11]. Besides, it is reported that caspase-11 is also
necessary for the activation of caspase-1. Caspase-11 can physically
interact with caspase-1 to promote its activation [12]. It has been
reported that type I interferon (IFN) could inhibit caspase-1 activity.
IFN-β signaling, via the transcription factor STAT1, represses the
activity of the NLRP1 and NLRP3 inflammasomes, thus suppressing
caspase-1-dependent IL-1β maturation [13]. Moreover, IFN-β could
also inhibit IL-1β transcription through STAT1 homodimers and
the ISGF3 complex which consists of STAT1, STAT2, and IRF9 [14].
In addition to caspase-1, one additional factor that can mediate IL-
1β processing and activation is proapoptotic enzyme caspase-8.
Caspase-8 is the major protease that cleaves pro-IL-1β during
infection with fungal pathogens. Fungal components activate
dectin-1 receptor signaling to induce a noncanonical CARD9–BCL-
10–MALT1–ASC–caspase-8 complex. Activated caspase-8 then
cleaves pro-IL-1β independent of the inflammasome complex [15].
The mammalian target of rapamycin (mTOR) is a conserved serine–
threonine kinase that is essential for the regulation of macrophage
inflammatory response.
Actually, mTOR complex consists of two complexes, rapamycinsensitive
mTORC1 and relatively rapamycin-insensitive mTORC2.
Tuberous sclerosis 1 (TSC1) and TSC2 are endogenous negative
regulators of mTORC1. Over-activation of mTORC1 through TSC1
deficiency could lead to decreased pro-IL-1β expression by inhibiting
C/EBPβ expression [4]. However, in the absence of TSC1/2, the
activity of RasGTPase is increased, which leads to the activation of
ERK [16] and then induces IL-1β transcription. Therefore, in TSC1
KO macrophage, the final IL-1β transcription is decreased as a result
of the antagonistic effect of these two pathways. The mTOR is a key
component to regulate the autophagy under cellular physiological
conditions and environmental stress. The inhibitory function of
mTORC1 in autophagy is well studied as mTORC1 activity reflects
cellular nutritional status. TSC1 deficient macrophages exhibit the
enhanced basal and mycobacterial infection-induced autophagy
[17]. In Yang’s paper, the activity of caspase-1 is suppressed by
over-activated mTORC1, which facilitates the decreased production
of active IL-1β [4]. However, the detailed molecular basis of mTORmediated
regulation of caspase-1 activation is yet unclear. It
would be interesting to investigate whether the altered autophagy
contributes to the reduced IL-1β expression in TSC1 deficient
macrophages.
References
- Dinarello CA (2011) Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood 117(14): 3720-3732.
- Hiscott J, J Marois, L Garoufails, MD Addario, A Roulston, et al. (1993) Characterization of a functional NF-kappa B site in the human interleukin 1 beta promoter: evidence for a positive autoregulatory loop. Mol Cell Biol 13(10): 6231-6240.
- Baldassare JJ, Y Bi, CJ Bellone (1999) The role of p38 mitogen-activated protein kinase in IL-1 beta transcription. J Immunol 162(9): 5367-5373.
- Yang T, L Zhu, Y Zhai, A zhao, LJ peng, et al. (2016) TSC1 controls IL-1beta expression in macrophages via mTORC1-dependent C/EBPbeta pathway. Cell Mol Immunol 13(5): 640-650.
- Yang Z (2000) NF-IL6 (C/EBPbeta ) vigorously activates il1b gene expression via a Spi-1 (PU.1) protein-protein tether. J Biol Chem 275(28): 21272-21277.
- Kaplan DH (2013) The IL-20 cytokine subfamily: bad guys in host defense? Nat Immunol 14(8): 774-775.
- Murray PJ, ST Smale (2012) Restraint of inflammatory signaling by interdependent strata of negative regulatory pathways. Nat Immunol 13(10): 916-924.
- Tannahill GM, Curtis AM, Adamik J, Palson EM, MCGettrick AF, et al. (2013) Succinate is an inflammatory signal that induces IL-1beta through HIF-1alpha. Nature 496(7444): 238-242.
- C Jefferies, C Wynne, R Higgs (2011) Antiviral TRIMs: friend or foe in autoimmune and autoinflammatory disease? Nat Rev Immunol 11(9): 617-625.
- Shi CS, Shenderov K, Hunag NN, Kabat J, Asab MA, et al. (2012) Activation of autophagy by inflammatory signals limits IL-1beta production by targeting ubiquitinated inflammasomes for destruction. Nat Immunol 13(3): 255-263.
- Shibutani ST, Saitoh T, Nowag H, Munz C (2015) Autophagy and autophagy-related proteins in the immune system. Nat Immunol 16(10): 1014-1024.
- Kayagaki N, Warming S, Lamkanfi M, Vande walle L, Louie S, et al. (2011) Non-canonical inflammasome activation targets caspase-11. Nature 479(7371): 117-121.
- Guarda G, Barun M, Staehli F, Tardivel A, Mattmann C, et al. (2011) Type I interferon inhibits interleukin-1 production and inflammasome activation. Immunity 34(2): 213-223.
- Castiglia V, Piersigilli A, Ebner F, Janos M, Goldmann O, et al. (2016) Type I Interferon Signaling Prevents IL-1beta-Driven Lethal Systemic Hyperinflammation during Invasive Bacterial Infection of Soft Tissue. Cell Host Microbe 19(3): 375-387.
- Gringhuis SI, Kaptein MT, Wevers BA, Theelan B, Boekhount T, et al. (2012) Dectin-1 is an extracellular pathogen sensor for the induction and processing of IL-1beta via a noncanonical caspase-8 inflammasome. Nat Immunol 13(3): 246-254.
- Zhu L, Yang T, Li L, Sun L, Hou Y, et al. (2014) TSC1 controls macrophage polarization to prevent inflammatory disease. Nat Commun 5: 4696.
- Pan H, XP Zhong, S Lee (2016) Sustained activation of mTORC1 in macrophages increases AMPKalpha-dependent autophagy to maintain cellular homeostasis. BMC Biochem 17(1): 14.
