info@biomedres.us
+1 (502) 904-2126
One Westbrook Corporate Center, Suite 300, Westchester, IL 60154, USA
Site Map
Received: June 12, 2017; Published: June 27, 2017
Corresponding author: Vincent van Ginneken, Blue green technologies, Ginkelseweg 2, 6866 DZ Heelsum, The Netherlands
DOI: 10.26717/BJSTR.2017.01.000151
Biomarkers of aging would give the true “biological age”, which may be different from the chronological age. Population perspectives regarding to aging, age-related diseases and life-expectancy of populations are extremely useful for e.g. governments and/or politicians. The problem with finding biomarkers of aging is that there are several theories of aging related to research areas like: oxidative stress, mitochondrial damage, telomeres shortage and cellular senescence, apoptosis and genetic predisposition. Although maximum lifespan would be a means of validating biomarkers of aging, it would not be a practical means for long-lived species such as humans because longitudinal studies would take far too much time. Ideally, biomarkers of aging should assay the biological process of aging and not a predisposition to disease.
Despite the fact that few biomarkers are found for aging some examples are given from Systems Biology: metabolomics, proteomics, genomics and lipidomics. While as somatic index Sarcopenia or “Nutritional Frailty” (≈nutritional stress), has been well documented for potential biomarkers of aging human brain research is underestimated. One target that has been looked at in this manuscript are biomarkers of brain aging. So far, brain function and age have proved too complex to produce reliable biomarkers. Here we present as a start material from the Netherlands Brain Bank, from a small cohort of elderly patients (>65 years) (all NIDDM diabetes patients) frequency distributions of the ultimate scope of the lifespan in neural tissue and some histopathological post mortem determined biomarkers such as Healthy (39.4%), mild- Alzheimer (32.8%), Dementia (13.5%) , Multiple Sclerosis (4.2%) several Dualistic Mix-forms (3.9%), Vascular Dementia (3.5%) and Parkinson (2.7%). Brain disease pattern were split up related to age and gender. Mix varieties and Vasculair Dementia are more common in the male brain while in the female brain mild-Alzheimer disease is more common. All the scores for brain somatic index (Control and diseased) are skewed to the left which is indicative for brain shrinkage of this elderly (>65+ years old) population. Brain diseases are not interrelated with Body Mass Index (BMI) in the elderly with type 2 diabetes so obesity is not the major cause for their morbidity.
Keywords: Biomarkers of aging; Elderly; Reductionism; Systems biology; Metabolomics; Proteomics; Genomics; Lipidomics; Somatic; Sarcopenia; Brain; Type 2 Diabetes; Mild-Alzheimer; Dementia; Multiple Sclerosis; Dualistic Mix-Forms; Vascular Dementia And Parkinson; Morbidity; Body Mass Index (Bmi)
Abbreviations: MLS: Maximum Life Span; ROS: Reactive Oxygen Species; HD: Huntington’s Diseases; HMDB: Human Metabolome Database; NHGRI: National Human Genome Research; GC-MS: Gas chromatography mass spectrometry; LC-MS: liquid chromatography mass spectrometry; MD-LCMS: multidimensional liquid chromatography mass spectrometry; NMR: Nuclear magnetic resonance; PTM: Post-translational modification; CR: Caloric Restriction; SPM30: senescence marker protein-30; SAA-1: Serum Amyloid protein A-1; NGS: Next Generation Sequencing ; BBB: Blood-Brain-Barrier ; VBM: Voxel-based-morphometry
Aging is a process that can affect almost all the systems in the body. With increasing age, physically and mentally healthy adults gradu¬ally become less fit and more vulnerable to illness and death. However, these changes happen at different rates in different people like is depicted in Figure 1 in the “global map of aging”. Conventional measures of aging are based on chronological age. But populations’ aging is not only about the percentage division of the different age groups in a certain population in a country or continent or during an era but also has more value if the population perspectives regarding to aging, age-related diseases and life-expectancy are encountered in the data. These “predicted age data” are more useful for e.g. governments and/or politicians, because they can anticipate on the expected age-related population structure for practical matters like elderly-care, public-health care, or retirement funding. The Maximum Life Span (MLS) is the upper boundary of life, the maximum numbers of years any human is known to have lived [1].
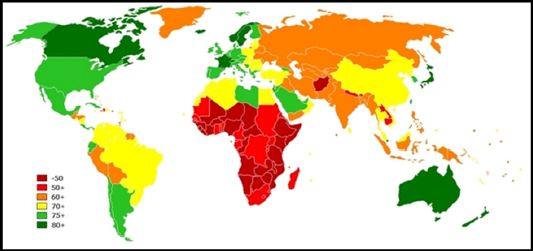
Figure 1: Maximum Lifespan (MLS) of the World population.
From Figure 1 we can see that the MLS of the Human Population World-Wide is the lowest in Africa ≈ 50 years or below, and the highest (75+-80+) in the industrialized countries like: Canada, USA, Argentina, Saudi Arabia, Libya, Europe, Japan and Australia. It is estimated that Human lifespan variation is mainly determined by environmental factors, whereas the genetic contribution is 25–30% and expected to be polygenic [2].
Lutz et al. [3] used new indicators of aging and all their measures indicate a continuous aging of the world’s population throughout the century. The proportion of the global population of 60+ years increases from 10% in 2000 to 21.8% in 2050 and then to 32.2% in 2100 [3]. When increases in life expectancy are taken into account these numbers are lower. The proportion of the global population of 60+ years old increases from 10.0% in 2000 to 21.8% in 2050 and then to 32.2% in 2100 [3]. These numbers are worldwide although there are large differences in the regional patterns of aging, example gratia for Western Europe the proportion 80+ is almost certain to increase significantly over the coming decades [3]. This brings us to Figure 2, where a world-wide estimation is given of the percent of the population at an age of 65+ in 2050 in comparison to 2000. The world is aging rapidly. The number of people aged 65 and over will double as a proportion of the global population, from 7% in 2000 to 16% in 2050. By then, there will be more elderly people than children (aged 0–14 years) in the population for the first time in human history. Chronic diseases, which affect older adults disproportionately, contribute to disability, diminish quality of life, and increased health- and long-term--care costs [4].
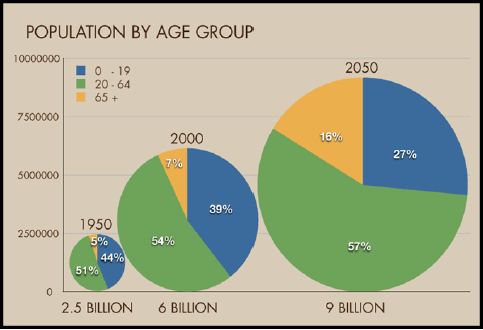
Figure 2: The aging World Population. (Source: Population Division of the Department of Economic and Social Affairs of the United Nations Secretariat, World Population Prospects: The 2006 Revision and World Urbanization Prospects: The 2005 Revision).
First we have to distinguish between chronological aging and biological (≈physiological) aging. Chronological aging is based on only one factor time. So when you are born in 2000 and we live now in 2010 your chronological age is 10: so current minus date of birth =chronological age. In other words, chronological age is a function of time and cannot be slowed, stopped, or accelerated. In contrast, in biological aging the physiological state of your body is considered. Our body becomes in general terms “out of Energy Homeostasis”. The existence of an organism depends on the continuous provision of energy for metabolic processes. The most important metabolic fuels are glucose and fatty acids [5]. The maintenance of narrow-controlled blood glucose concentrations (glucose homeostasis) is central for a constant provision of glucose to the brain [6,7]. Glucose homeostasis is a physiologically well-balanced mechanism depending on three coordinated and simultaneously ongoing processes involving insulin secretion by the pancreas, hepatic glucose output and glucose uptake by splanchic (liver and gut) and peripheral tissues (muscle and fat). During aging and obesity animals/humans are “Out of Energy Homeostasis”. Humans over 70 year of age often lose weight. This appears to be due to physiological anorexia of aging as well as a loss of lean mass (Sarcopenia or “nutritional frailty ≈ stress” [8]) and, to a lesser extent, fat mass. Central is the term “Nutritional Frailty” (≈nutritional stress), which refers to the disability that occurs in old age owing to rapid, unintentional loss of body weight and loss of lean body mass. This loss of muscle mass and strength, contributes to functional impairment [9] (Figure 3).
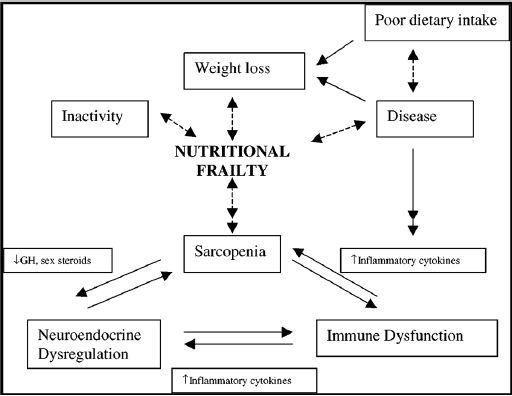
Figure 3: Dimensions of nutritional frailty in the elderly (Source: [8]). GH=growth hormone.
There are several underlying mechanisms at Sacropenia including intrinsic changes in the muscle and central nervous system, endocrine and lifestyle factors. For muscle this leads to age related deterioration of muscle contractility at the whole muscle level and at the single fiber level [10]. Very recently via a reductionistic approach the current knowledge on biological markers for sarcopenia was summarized, including a critical appraisal of traditional procedures for biomarker development in the field of muscle aging. As an alternative approach using a more systems biology approach the researchers illustrate the potential advantages of biomarker discovery procedures based on multivariate methodologies. Relevant examples of multidimensional biomarker modeling are provided with an emphasis on its clinical and research application [11]. Lara et al. [12] mention an array of biomarkers in the following domains:
i. Physiology: related to muscle-, lung and glucose metabolism;
ii. Memory: processing speed and cognitive function;
iii. Endocrine: HPA-axis, sex hormones, growth hormone;
iv. Immune system;
Inflammatory factors [12]. However, Critics also argue that aging doesn’t occur at a single rate in several tissues, and therefore the rate of aging can’t be measured. Different body functions can change at different rates over time (Figures 4 & 5). If we know the mechanisms of aging we can targeted search for biomarkers.

Figure 4: Increase of age-related diseases in Humans during aging.
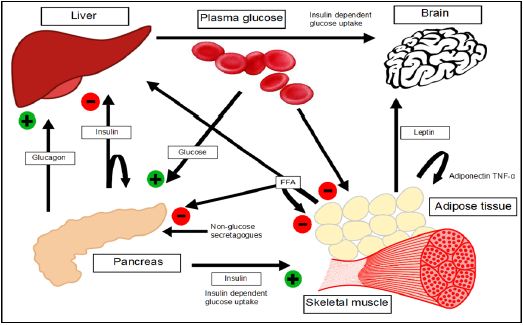
Figure 5: Cross-talk between tissues in the regulation of glucose metabolism. Insulin is secreted from the β-cells of the pancreas in response to elevations in plasma glucose e.g. after a meal. The hormone decreases glucose production from the liver, and increases glucose uptake, utilization and storage in fat and muscle. Main target is to fuel the brain continuously with glucose. The fat cell is important in metabolic regulation, releasing FFAs that reduce glucose uptake in muscle, insulin secretion from the β-cell, and increase glucose production from the liver. The fat cell can also secrete adopokines such as leptin, adiponectin, and TNF, which regulate food intake, energy expenditure and insulin sensitivity.
A biomarker is a characteristic that can be objectively measured as an indicator of normal biological processes, pathogenic processes or a pharmacological response to a therapeutic intervention. These are used for many purposes including disease diagnosis and prognosis, prediction and assessment of treatment response. Biomarkers can be characteristic biological properties or molecules that can be detected and measured in parts of the body like the blood or tissue. They may indicate either normal or diseased processes in the body. Biomarkers can be specific cells, molecules, or genes & gene products, enzymes, or hormones.The problem with finding biomarkers of aging is that there are several theories of aging given below.
I. Oxidative Stress: From a whole animal level “rate of living hypothesis” explain the observation that animals with higher metabolic rates have shorter lifespans due to the role of reactive oxygen species (ROS) in aging [13]. From a cellular level the free radical theory of aging was first proposed by Denman Harman in the 1950s [14]. There is now substantial evidence that supports that aging is associated with, if not the consequence of, free radical damage by various endogenous Reactive Oxygen Species [13,15].
II. Memory: processing speed and cognitive fuMitochondria and Aging: Mitochondria are both producers and targets of oxidative stress; this fact forms the basis for “the mitochondrial theory of aging” [16]. It has been proposed that accumulation of somatic mutations of mitochondrial DNA, induced by exposure to reactive oxygen species (ROS) generated within mitochondria, leads to errors in the mitochondrial DNA-encoded polypeptides and subsequent defective electron transfer activity and oxidative phosphorylation. Such respiratory chain defects lead to increased reactive oxygen species production (Figure 6).
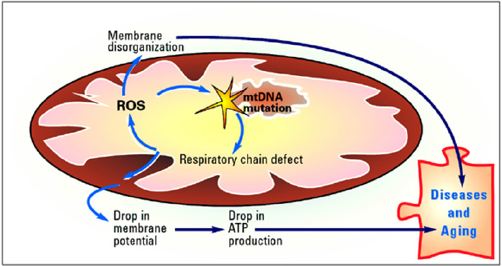
Figure 6: One of the features of the modern westernized welfare diseases –but also during aging- is mitochondrial dysfunction. Some possible causes are: i. Mitochondrial DNA (mtDNA) mutations caused by free radical damage (ROS-escape). ii. Krebs’’ cycle decreased efficiency due to inadequate Krebs’ cycle intermediates. iii. Respiratory chain defect due to enzyme and/or substrate alterations. iv. Membrane disorganization and loss of fluidity.
III. Telomeres and Cellular Senescence: In culture, diploid cells exhibit a limited proliferative potential. After a finite number of divisions, primary cell cultures enter a state of replicative senescence. This number of divisions, known as the Hay flick limit [17], has been postulated to determine the maximum lifespan of an organism [18]. One explanation for cells reaching this limit arises from telomeres, the repetitive DNA sequences at the end of linear DNA. Telomeres shorten slightly each time the cell divides. Depletion of telomeric DNA prohibits further cell division.
IV. Apoptosis: Aging is associated with dysregulation of apoptosis [19], and overall, it has been suggested that aging is mostly associated with up-regulation of apoptosis [20]. It is not clear whether age-related dysregulation of apoptosis is the result of genetic programming or stochastic aging processes such as oxidative stress [18].
V. Genetic Mechanisms for Aging: The role of genetically programmed aging is still controversial [21]. Evidence for a primary role for genetic programming includes the observations that the lifespan of a given species is relatively fixed and human aging has a hereditary component. In addition [22] used a gene expression database for mouse (AGEMAP) and found the important observation that aging within an organism was strongly organ/tissue dependent (splitting up to aging patterns for: neural, vascular tissue and steroid response tissue [22].
Figure 7: The complexity of the Human metabolism with its complex of biochemistry and the comprehensive metabolic chart of living organisms (medical biochemistry) and the interaction between the different metabolic routes and biochemical pathways in healthy organisms or during disease [5,6,7].
Presently researchers are looking for a more complete understanding of the mechanisms of aging, to answer questions about the biological processes that account for an inevitable decline in physical vitality. In other words, is aging a single process that affects many of the body’s systems, or are separate processes going on within each system? Currently, the only way to test interventions that are aimed at extending life is to conduct studies that follow subjects to the end of their lives. This can take an imprac¬tically long time for humans. What’s needed is biomarkers of the aging process that could help determine a person’s life expectancy, making it unnecessary to wait many years for the results of studies. Ideally, there would be a set of these biomarkers that would identify biological age. These could be used to test whether a behavior (such as exercise), drug, or dietary additive slowed down the aging process. If a set of biomarkers of aging were identified, it would also have the effect of demonstrating that there actually is an underlying mechanism of aging that coordi¬nates changes across the body’s systems (Figure 7).
Reductionism can either mean
a) An approach to understanding the nature of complex things by reducing them to the interaction of their parts, or to simpler or more fundamental things or
b) A philosophical position that a complex system is nothing but the sum of its parts, and that the account of it can be reduced to accounts of individual constituents. In contrast, Systems Biology has a holistic view and focuses on complex interactions in biological systems, claiming that it uses a new perspective.
The Reductionistic vs. Systems Biology approach is related to a discipline example gratia reductionism is propagated in sciences like Chemical Engineering [23], Biochemistry and Biophysics [24], while Systems Biology is propagated in Sciences like Biology, Pathobiology [25]. The major “drive” and interest for a Systems Biology approach is linked to the progress in collecting tremendous experimental datasets and at the system-level understanding of native biological and pathological systems to provide potential therapeutic targets [26]. Medicine is at the edge, clinical medicine focuses on the parts and the Systems Biology on the systems [25,26]. While the “traditional” approach in medicine is based on four principles:
a)The focus on a singular, dominant factor which is the deeply rooted belief that each disease has a potential singular target for medical treatment. For example, an infection has to be treated with antibiotics but the underlying mechanism example gratia an impaired immune system is not treated.
b)Emphasis on homeostasis. This is based on two principles:
i. Homeostasis needs to be maintained by placing a deviating physiological parameter (example gratia high blood pressure) within its physiological range,
ii. Because reductionism often disregards the dynamic interaction between parts, the system is often depicted as a collection of static components.
c) Inexact risk modification. The often in medical epidemiology “one risk-factor to one-disease” approach has certain limitations.
d) Additive treatments. This is characterized by a reductionistic approach: one risk to one-disease analysis and the inability example gratia for age-related diseases with multiple risk factors and calculate their collective influences [26].
The need to make sense of complex genetic interactions has led some researchers from a component-level to system-level perspective. In understanding the difficulty of complex biological systems research has to be directed to the components of a biological system, their interactions and the behaviors and properties of the whole system. This is the basis for a Systems Biology approach. The progress made in this new research area is also related to the progress made the last decade in other research areas like: Molecular Biology, Computational Science, Statistics, Chemistry, and Mathematics [26]. Also for biomarkers of aging recently by [27] Systems biology was warranted because it aims to integrate data from different experiments to gain an understanding of the system as a whole rather than focusing on individual factors. Furthermore, it thus allows deeper insights into the mechanisms of complex traits, which are caused by the joint influence of several, interacting changes in the biological system. The final target would be the need for further developments in this area to improve epidemiologic investigations [27].|
Via a reductionistic approach animal mouse mainly with genetically engineered mice were used to get insight which genes are actually mutated in human disease. By eliminating these genes (knock-out) or inserting mutations into specific genes, creating transgenic animals, (knock-in) some mechanisms of the aging process can be elucidated and possibly via these approach biomarkers for senescence can be found.
-Example knock-in mice model: In order to give an answer of the point mutations and deletions of mitochondrial DNA (mtDNA) accumulate in a variety of tissues during aging in humans and this increase in somatic mtDNA mutations is associated with reduced lifespan [28]. Trifunovic et al. [28] created a homozygous knockin mice that expresses a proof-reading-deficient version of POlgA, the nucleus-encoded catalytic subunit of mtDNA polymerase. Their study with the knock-in mice strain shows that the animals develop an mtDNA mutator phenotype with a threefold to fivefold increase in the levels of point mutations, as well as increased amounts of deleted mtDNA. This increase in somatic mtDNA mutations is associated with reduced lifespan and premature onset of agingrelated phenotypes. Their results clearly provide a causative link between mtDNA mutations and aging phenotypes in mammals [28].
Another example is the creation of Hdh CAG(150) homozygous knock-in mice to study Huntington’s diseases (HD), an incurable and fatal neurodegenerative disorder characterized by loss of stratial mediums spiny neurons which finally lead to muscle disorders [29]. In this Hdh CAG(150) homozygous knock-in mice model the HD-triggered gene expression phenotype is consistent with the beginnings of a transition from fast-twitch to slow-twich muscle fiber types. In addition, metabolic adaptations similar to those induced by diabetes or fasting (=> Caloric Restriction) are also present but neither metabolic disorder can explain the full phenotype of HD muscle.
-Example knock-out mice model: A mouse strain knockout for the senescence marker protein-30 (SPM-30) had after three months an increased mortality rate, and many alterations in the liver like, many empty vacuoles, abnormally enlarged mitochondria, increased hepatic triacylglycerol-, cholesterol and phospholipid levels compared to wild mice. The researchers suggest that the SPM-30 gene has an anti-apoptotic (note: apoptosis=programmed cell death) function with a wide spectrum and that this strain might be a useful tool for aging and biological monitoring [30]. However [31] critically noted –using these transgenic mouse models- that several questions still need to be addressed:
a) Most of the studies using the knockout and transgenic approach do not take into account the effect of oxidative stress on development. It is possible that the genetic manipulations are affecting development and not necessarily the aging process,
b) Our understanding of the mitochondrial biology is far from complete and it remains to be seen if any of the mouse proteins in turnover and maintenance of mitochondria affect the lifespan of higher organisms like human beings,
c) Results between mouse strains are not always consistent and uniform which possibly can be ascribed to differences in genetic background between the different mouse strains [31].
Other researchers also raised the question if we can extrapolate the results of mouse models to the human situation. Maegawa et al. [32] studied DNA methylation in normal colon tissues in young and old mice and found evidence of changes affecting over 20 percent of genes in the process of aging. DNA methylation is an epigenetic event that affects cell function by altering gene expression. This agerelated methylation instability is tissue-specific but the researchers questioned if this age-related DNA methylation phenomenon is specific to human tissues. So the question arises if a mouse model is suitable for the human situation [32].
There are two major critical points related to the reductionistic approach in order to detect major risk-associated genes coding for proteins involved in aging:
i. Mice are mammals but besides large (genetically caused?) differences between strains the question can be posed if the results of mouse studies can be extrapolated to the human situation.
ii. Mice are mammals but besides large (genetically caused?) differences between strains the question can be posed if the results of mouse studies can be extrapolated to the human situation.
So the reductionistic approach with transgenic/knock-out strategies has in most cases failed to unravel candidate genes (≈potential biomarkers), in the “garden variety” of the genetics of senescence (assuming there is such an entity). Therefore a “Systems Biology” approach is clearly warranted. The “Systems Biology” approach aims to understand phenotypic variation to assemble comprehensive data and models of cellular organization and biochemical function, and to elucidate interactions and pathways [33].
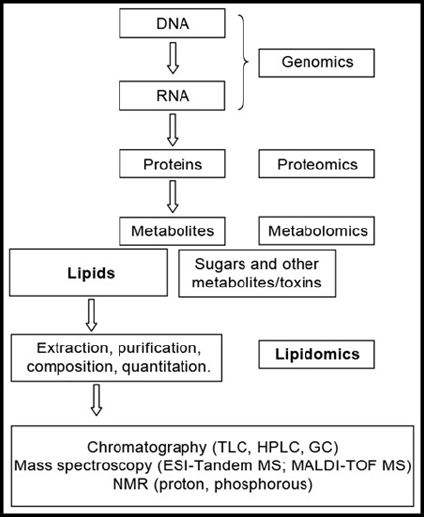
Figure 8: Overview of the “omics” technologies (genomics, transcriptomics, proteomics, metabolomics and lipidomics) at several levels in the organism to elucidate the mechanism in a certain disease, or elucidate a biochemical pathway or mechanism or developing predictive tools like “biomarkers” which can be applied in a personalized medical treatment.
Figure 9: One of the challenges of “omics” approach in Human metabolism is determining the significance of metabolomic changes. Finding changes in metabolite profile is not a problem but it is figuring out what these changes mean. This can eventually lead to novel safe biomarkers that can be used in the assessment of new intensive treatments for aging and in clinical use in combining the diagnostics with the patient therapy.
In principle metabolomics can provide certain advantages relative to other “omics” technologies (genomics, transcriptomics, proteomics) in human aging like is reviewed by [34] (Figures 8 & 9).
I. Estimates vary, but e.g. in the Human Metabolome Database (HMDB)-Canada [35], currently lists ~ 6,500 discrete small molecule metabolites, significantly less than the estimate of 25,000 genes, 100,000 transcripts, and 1,000,000 proteins.
II. Metabolomics measures chemical phenotypes that are the net result of genomic, transcriptomic, and proteomic variability, therefore providing the most integrated profile of biological status.
III. Metabolomics is in theory a precise tool for discerning mechanisms of action (see further “personalized medical treatment “ [36] and possible toxicological effects of drug therapies or in case of starvation of malnutrition the identification of novel and safety biomarkers that can be used in the assessment of new intensive treatments e.g. the development of new “functional foods” [37].
IV. An important area in which metabolomics has great potential is the discovery of biomarkers related to metabolic processes or age related diseases [38-41]. This is another perception as biomarkers of aging.
Over the past century, traditionally medical care has centered on standards of care based on epidemiological studies of large cohorts. However, large cohort studies do not take into account the genetic variability of individuals within a population. E.g. at the level of genomics, according the National Human Genome Research (NHGRI) about 99.9 percent of the DNA sequence is identical in all people, but the 0.1 percent difference is critical because it represents the genetic variations in the susceptibility of a person for getting a disease, the severity of the disease, and what is the impact of the medical/therapeutic treatment. Personalized medical treatment seeks to provide an objective basis for the consideration of such individual differences. It is often defined as “the right treatment for the right person at the right time” [42]. “Traditionally, personalized medicine has been limited to the consideration of a patient’s family history, social circumstances, environment and behaviors in tailoring individual care. Nowadays all the information generated by genomics (studying variations in genes that cause disease), proteomics (seeking for abnormal protein patterns) and metabolomics (mining for abnormal metabolite patterns) collected by the application of new measurement tools (GC-MS: Gas chromatography mass spectrometry; LC-MS: liquid chromatography mass spectrometry; MD-LCMS: multidimensional liquid chromatography mass spectrometry; NMR: Nuclear magnetic resonance; PTM: Post-translational modification [36] was one of the preconditions for the strategy of a personalized medical approach. The other precondition was the development of techniques in the processing of data by Bioinformatic-science, computation and modeling in combination with the development of new statistical techniques for application in biomedical research to evaluate the tremendous amount of datasets [43,44]. These trends during the last decade in Systems- Biology gave an enormous boost to this research area and we are close to a personalized diagnostic “Industry” [45]. Examining biomarkers for aging in personalized medicine would provide a scientific baseline for clinical trials of anti-aging medicines, which is currently impossible to determine given the lengthy lifetime of human beings. The technology would also provide a means of determining whether an individual is aging faster or slower than would normally be expected (Figure 10).
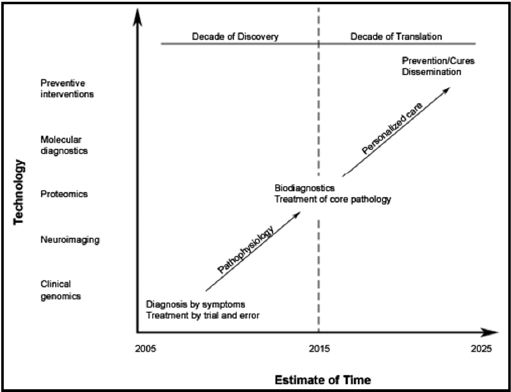
Figure 10: Perspective view how medical science based on an “omics” approach will develop towards a personalized medical treatment.
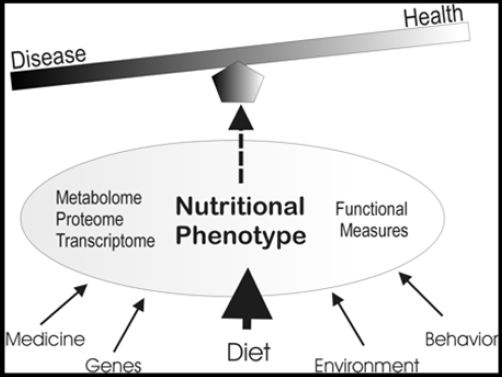
Figure 11: The factors which determine the balance between healthy or diseased.
A particular area of interest of Metabolomics-based System Biology, will be identification of novel safety biomarkers that can be used in the assessment of new intensive treatments both during their discovery phase and in clinical use e.g. to study age-related diseases in combining the diagnostics with the patient therapy [36]. Recent technological advances in metabolomics [43,44] and lipidomics [46] can potentially make a real contribution in increasing efficacy of drug development pipelines [47]. Metabolomics is a discipline dedicated to the systematic study of small molecules (ie, metabolites) in cells, tissues, and different bio-fluids. Metabolite levels can be regarded as amplified responses of biological systems to genetic or environmental changes [47]. From Figure 11 we can see that the balance between “diseased” or “healthy” is determined by factors like on one hand: environment, behavior and diet and for a diseased person by genetics and medical treatment. As a subgenre of bio-gerontology, biomarker research has developed a thoroughly tarnished reputation. The difficulties have several root causes:<
A. Mathematical problems because the development of techniques in the processing of data by Bioinformatics Science, computation and modeling in combination with the development of new statistical techniques for application in biomedical research to evaluate the tremendous amount of datasets were at that time not developed at that time.
B. Philosophical problems, typified by the objection that biomarkers cannot measure aging because there is no such thing as aging.
C. Research was less fruitful than governments and sponsors hoped.
D. The “Missouri syndrome”: An attitude of sceptic: ‘I just believe it when I see it’.
E. Overlap between aging and disease: both the aging process and diseases can cause changes in the body, which affect lifespan. It has been difficult, however, to separate out the aging process from conditions that precede the onset of disease, as growing older predisposes a person to many illnesses.
F. Some age-related changes cause no harm, while others do—and medical science doesn’t always know which is which. This makes it more difficult to pinpoint exactly what to look for.
G. Critics also argue that aging doesn’t occur at a single rate, and therefore the rate of aging can’t be measured.
H. Different body functions can change at different rates over time.
According to the [48] a true Biomarker for Human aging must satisfy the following criteria:
I. The marker must predict the rate of aging and be a better predictor of life span than chronological age.
II. It must be able to be tested on a regular basis
III. It must work both for humans and other species, such as laboratory animals.
IV. There is support from human clinical assessment and complementary research studies.
V. The studies are based on a significant representative sample.
VI. The result is a clear association with aging.
VII. A relatively narrow standard deviation is present.
VIII. Miller [49] added the following three criteria: It should predict the outcome of a wide range of age-sensitive tests in multiple physiological domains, in an age-coherent way, and do so better than chronological age.
IX. It should predict remaining longevity at an age which 90% of the population is still alive [49].
X. Its measurement should not alter either life expectancy of the outcome of subsequent tests of other age-sensitive traits [49].
In this respect molecular mediators in blood plasma are important to study because they can serve as biomarkers to trace people vulnerable and susceptible for the pathogenesis or aging.
The process to find a proper biomarker is underestimated and has to follow a long route:
a) The Discovery phase,
b) Qualification phase one, which encompasses:
i. Identification of the biomarker,
ii. Validation analysis,
iii. Proof of the biomarker in a new study,
b) Qualification phase two, which encompasses:
i. Study precision,
ii. Study robustness,
iii. Study limitation,
iv. Test biomarker in wider population, Finally the application phase in the population is reached [50].
There are already some studies in animals or humans which gave potential biomarker candidates for aging. Most of them are at the level of
A. Metabolomics (measuring metabolites),
B. Proteomics (measuring proteins),
C. Genomics (microarray profiling of gene expression) or
D. Lipidomics (measuring lipids).
I. Metabolomics: Huffman et al. [51] studied the effect of exercise and Caloric Restriction (CR) at the level of metabolomics studying: weight, serum IGF-1, fasting insulin and heat shock proteins. In CR-animals insulin and IGF-1 were the lowest. In contrast, the exercise experiencing mice were leaner, and had a slightly lower serum IGF-1. The researchers conclude that exercise cannot fully mimic the hormonal and/or metabolic responses to CR. In contrast, very recently a list of 19 human biomarkers of aging was tabulated grouped by function by [52] and enlisted in Table 1.
Table 1: List of the 19 biomarkers used to define signatures. The biomarkers are grouped by functions. Biomarkers with generally increasing values with older age are labeled with an arrow pointing up, while biomarkers that generally decrease with older age are labeled with an arrow pointing down (Source: [52]).
II. Proteomics: Maruyama et al. [53] studied a mouse strain lacking senescence marker protein-30 (SPM30). In this knockout model mortality rate was increased but also many hepatic lipid compound were increased. The researchers concluded that SPM30 might be a protein with an anti-apoptosis function and that eliminating the gene for SPM30 would result in physiological changes observed during senescence. Mi et al. [54] performed a study at age-related sub-proteomic analysis of peroxisomal proteins of mouse liver and kidney in cellular aging. The results indicate that peroxisomal proteins show a tissue-specific functional response to aging. This response is probably dependent on their differential regeneration capacity.
As a consequence cellular effects like alteration of fatty acid metabolism that could alert membrane protein function, increase of the oxidative stress and contribute to decline in bile salt synthesis could be affected. The researchers conclude that the ability to detect age-related variations in the peroxisomal proteome can help in the search for reliable and valid aging biomarkers. In male C57BL/6J mice (N=8) model, proteomics related biomarkers of aging were evaluated in a longitudinal study trough 2, 4, 8, 16 and 19 months of age. Using among else two-dimensional gel techniques the following proteins for aging were measured. For example, 23 isoforms of albumin, seven isoforms of transthyretin (TTR), four isoforms of haptoglobin (Hp), and three isoforms of immunoglobulin kappa chain (Ig kappa) were identified. Serum amyloid protein A-1 (SAA- 1), and peroxiredoxin-2 (Prx-2) were identified as single spots in this mouse model [55].
III. Genomics: Weindruch et al. [56] studied molecular events associated with aging and its retardation by caloric restriction (CR) were studied in mouse tissues using high-density oligonucleotide microarrays to define transcriptional patterns. Transcriptional patterns of muscle of CR animals suggest that CR retards the aging process by causing a metabolic shift towards increased protein turnover and decreased macromolecular damage. In addition, [57] also used high-density oligonucleotide microarrays to define transcriptional patterns. Transcriptional patterns of muscle of CR animals suggest that CR retards the aging process by reducing endogenous damage and by inducing metabolic shifts associated with specific transcriptional profiles. The same researchers used in the same year, DNA microarrays to examine transcriptional alterations associated with aging in skeletal muscle and the heart in a mouse model during CR. CR retards the aging process by reducing endogenous damage and by inducing metabolic shifts associated with specific transcriptional profiles [58]. A few years later, [59] also used DNA microarrays to identify panels of transcriptional markers of aging in heart- and brain tissue of mice strains. In the heart five genes were studied throughout the mouse lifespan:
i. complement component 4,
ii. chemokine ligand 14,
iii. component of Sp100-rs,
iv. phenylalanine hydroxylase and
v. SRC family associated phosphoprotein-2. For the brain a similar analysis was performed with the
following genes:
i. 1q (alpha polypeptide),
ii. Complement component 4,
iii. Plysozyme structural,
iv. Glial fibrillary acidic protein and
v. Cathepsin S. Caloric Restriction inhibited age-related expression of these genes in both tissues.
The antioxidants Lycopene, reveratrol, acetyl-L-carnitine and Tempol were as effective as CR in the heart while α-lipoic acid and coenzyme Q10 were as effective as CR in the brain. The researchers conclude that transcriptional biomarkers of aging in mice can be used to estimate the efficacy of aging interventions on a tissuespecific basis. Human genomic research is presently progressively being integrated with biomarker studies on aging, including the application of (noninvasive) deep phenotyping and omics data. Explorative and candidate gene studies of the human genome by genetic, transcriptomic, and epigenomic approaches have resulted in the identification of a limited number of interesting positive linkage regions, genes, and pathways that contribute to lifespan variation [2]. The possibilities to further exploit these findings are rapidly increasing through the use of novel technologies, such as next-generation sequencing. However, results didn’t pan out as expected. Exploration of the common variation in the genome did not explain much of the variation in the rate of aging and longevity, researchers in the field have only recently started to investigate the contribution of rare genetic variants to these traits. The increased throughput and decreased costs of next generation sequencing (NGS) have resulted in a great deal of novel applications for sequencing sets of candidate genes, whole exomes, and whole genomes of individuals [60].
IV. Lipidomics: In this respect, one target that has been looked at is the central nervous system. Computerized tomography (CT) scans can be used to look for changes in the brain that may serve as biomarkers of aging. For example, the brain shrinks with age. This does not mean that brain cells necessarily die, but rather that they become smaller in size and volume. Some research has been performed by Voxel-based-morphometry (VBM) for whole-brain as unbiased technique for characterizing regional cerebral volume and tissue concentration differences in structural magnetic resonance images [61]. Brain shrinkage was found in grey matter of mainly males with increasing age [61]. So far, brain function and age have proved too complex to produce reliable biomarkers.
Throughout the last 200 years, the scientific community has meticulously characterized and categorized the pathological processes of diseases of aging with major emphasis on our hormonal glands in the body, followed by our organs in the body and finally followed by neural tissue [62]. A definition for “Dementia”, is a progressive decline in two or more cognitive domains that is severe enough to interfere with the performance of everyday activities [63]. Deficits in cognitive function contribute to impaired functional status [64]. The perception of old age as a pathology unto itself persisted throughout the Greco-Roman period [65]. The term “senility” was synonymous with “dementia” or “imbecility”, despite its origin from the word “senile,” or “aged” in Latin. We know now that, while the aged human body no doubt falls victim to long-term wear and tear, there are numerous examples of healthy human aging and that diseases of aging are not inevitable [66-68] (Figures 12 -14).
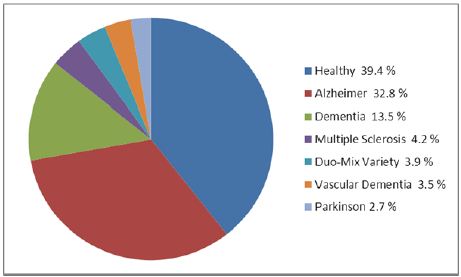
Figure 12: Nervous tissue is the least vulnerable to aging.
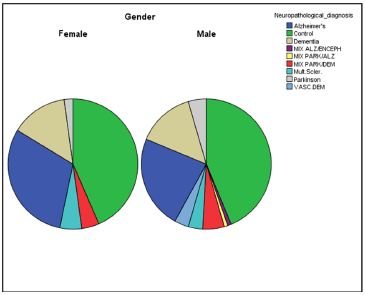
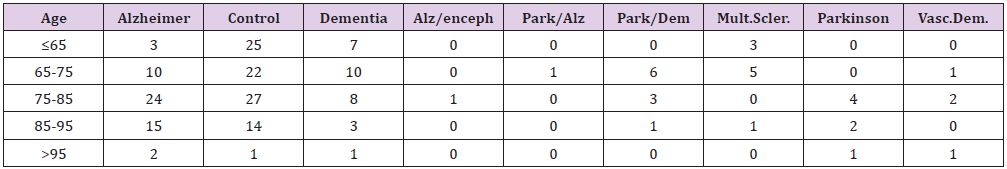
Figure 13: Our basic cohort consisted of 306 patients with diagnosed diabetes of which 259 patients (84.6%) were elderly patients (>65 year). In addition for other criteria (annex 1) patients were excluded giving a total cohort of 204 patients. Based on microscopic classification based on histological pathophysiological characteristics we quantified in this cohort of elderly Noninsulin-dependent diabetes mellitus (NIDDM) patients: 39.4% healthy (Cogroup); 32.8% Alzheimer (mild form); 13.5% “real” Dementia; 4.2% Multiple Sclerosis; 3.9% several dualistic mix-forms; 3.5% Vascular Dementia and 2.7% Parkinson.
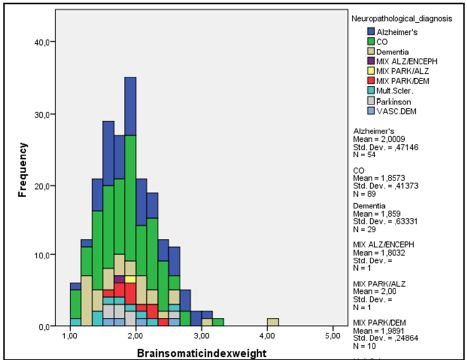
Figure 14: Frequency distribution in a cohort (n=204), split up to gender of the Netherlands Brain Bank of morbidity of “brain diseases” as ultimate biomarkers of aging. (Courtesy: Evert de Vries).
However it seems that the human brain and in general neural tissue is the least vulnerable to aging [62]. Interestingly, in an animal mouse model using a gene expression database (AGEMAP: Atlas of Gene Expression in Mouse Aging Project).In this gene expression database (AGEMAP), 8,932 genes in 16 tissues could be monitored as a function of age. Surprisingly, the aging in an organism was strongly organ/tissue dependent. The researchers found that organs/tissues could be classified into one of three aging processes:
i. A pattern common to neural tissue,
ii. A pattern for vascular tissue,
iii. A pattern of steroid-responsive tissues [22].
The study of [22] with an aging mouse model is very important with the perception it clearly demonstrates the aging process in time. At first the hormonal glands, followed by the organs in the body and finally the brain.
Evidence from well-planned, representative epidemiological surveys is scarce in many regions. Ferri et al. [69] estimated that 24•3 million people have dementia today, with 4•6 million new cases of dementia every year (one new case every 7 seconds). The number of people affected will double every 20 years to 81•1 million by 2040. Most people with dementia live in developing countries (60% in 2001, rising to 71% by 2040). The number of people affected will be over 100 million by 2050. Therefore it is important to elucidate the mechanisms behind “brain-diseases” for which its morbidity in an will be further outlined based on unique post mortem brain material at a cohort of around 300 patients > 65 years of the “Netherlands Brain Bank” will be given below as ultimate “biomarkers of aging” at the extreme end of the Maximal Lifespan. We believe that the detailed estimates in this review/ research manuscript constitute the best currently available basis for policymaking, planning, and allocation of health and welfare resources.
Notify the mix varieties and vasculair dementia are more common in the male brain while in the female brain mild-Alzheimer disease is more common. The following three mix varieties were observed in the male >65+ years cohort: mix Alzheimer/ Encephalopathy; mix Parkinson/Alzheimer and mix Parkinson/ Dementia. So more than one type of dementia may exist in the same person.
The lipid component rich brain is not always protected by the blood- brain-barrier (BBB) which was demonstrated for Type-2 diabetes (DM2) patients which are at increased risk of cognitive impairment. Micro-vascular damage which has clearly been demonstrated in DM2 patients in small arterioles to leak resulting in retinal and renal complications might also cause cerebral damage by the same small arterioles to leak in the head region resulting in a loss of blood-brain-barrier (BBB) integrity [70]. The study of [70] shows for DM2 patients that micro-vascular complications has as consequence the loss of the blood-brain-barrier (BBB) integrity. Due to leakage of small arterioles start the brain constitutes not anymore a mechanism of protecting it from self-destruction and maintaining its integrity. Due to catabolism under these conditions mainly the brain lipids and lipid-protein interactions within membranes for energy production the brain volume will diminish and the process of dementia will further speed up resulting in deteriorating and impaired cognitive functions. Lipids play a key role in determining membrane fluidity, and changes in lipid composition have been reported to alter cellular functions [71,72].
As a consequence membrane lipid composition appears to influence the state of mental health and is likely to exert its effects through lipid-protein interactions within the membrane. Such interactions may include effects upon neurotransmitter release and reuptake, and membrane receptors [73]. We hypothesize that in case of vascular dementia a similar mechanism as [70] described for DM2 patients will occur in patients with vascular dementia resulting in case of a deteriorating pathogenesis with as consequence a direct negative correlation with brain volume. In other words with decreased protective function of the BBB in patients with vascular dementia more micro-vascular complications will result in leakage of blood out of small arterioles which will use the lipid and membrane lipid-protein brain structures as substrates for bodily catabolism. The final consequence is that the brain volume will decrease with progress of the disease depending on unwanted “cross-talk” between the brain and the body compartment due to leakage of blood and catabolize brain components. Brain volume decrease is a rough anatomical parameter but is directly related to cognitive brain processes and behavioral functions directly related to the quality of life of the patient and its dependence on hospitalizing personal. This ultimately has a total effect for the cognitive function of patient, mood, memory, self-awareness, character changes, recognition of relatives: all processes more or less related to molecular processes in the lipid membrane, receptor proteins, and neurotransmitters.
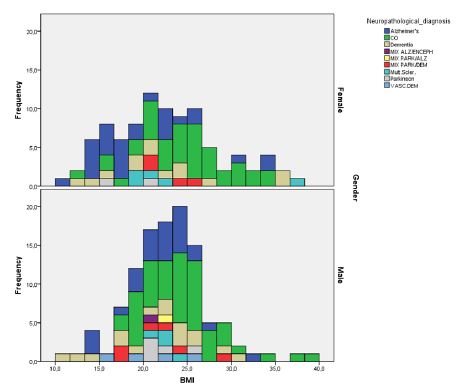
Figure 15: Frequency distribution for the braindomaticindex, which index is defined as brain weight divided by the somatic weight x 100% of this elderly (>65+ years old) population (n=204) (Courtesy: Evert de Vries).
From Figure 15 we very clearly can see that all the scores for brain somatic index (Control and diseased) are skewed to the left which is indicative for brain shrinkage of this elderly (>65+ years old) population. This observation has also been performed based on another approach of Voxel-based-morphometry (VBM) for wholebrain as unbiased technique for characterizing regional cerebral volume and tissue concentration differences in structural magnetic resonance images. Good et al. [61] described an optimized method of VBM to examine the effects of age on grey and white matter and CSF in 465 normal adults. Global grey matter volume decreased linearly with age, with a significantly steeper decline in males [61].
Figure 16: Normal distribution curves of Body Mass Index of a small cohort (n=204) of the Netherlands Brain Bank for which histopathological the “brain diseases” post mortem were determined. Gauss-curves are split up to gender. Note: The BMI is defined as the body mass divided by the square of the body height, and is universally expressed in units of kg/m2, resulting from mass in kilograms and height in meters. (Courtesy: Evert de Vries).
Symmetry is the most important characteristic of the normal distribution. As one knows each normal distribution lies symmetrically around the mean. Figure 16 tells us that most patients in this small cohort of 204 patients are close an average value of the Body Mass Index (BMI) around 23 while relatively fewer cases tend to one extreme or the other. The standard deviation is a statistic parameter reflecting you how tightly all the various examples are clustered around the mean in a set of data. Here we observe especially in case of the gender females the bell curve is relatively flat, that is a reflection we have a relatively large standard deviation. Among all anthropometric measurements, the BMI represents the easier and most frequently used index to identify subjects at risk for under- or over-nutrition.
The value of the BMI is generally considered to be as a measurement of fatness, while it also gives information about fat-free mass. In the elderly, fat mass increases whereas fat-free mass decreases [74]. The same adult BMI value corresponds to a more fatty body composition in the elderly. Thus the BMI has to be differently interpreted for elderly subjects. However from our observations we conclude, in general terms spoken some patients (especially females) have a BMI > 30 and are obese up to even a BMI of 40, healthy and diseased. On the other side of the scale, undernutrition is a well-known predictor of mortality. Some patients, especially Alzheimer and dementia patients (both gender) even have a BMI in the range of 10-15, which value can been characterized as heavenly undernutrition. In earlier work we mentioned for males a limit of survival with a BMI of 13 (STD 8.7%) while for females this cut-off value was around 11 (STD 14%) [75]. At this stage one can start treatment with anodyne medicine like morphine for palliative treatment and “passive euthanasia” can be from medical perception been discussed [76] (Figure 17). In general terms spoken, brain diseases are not interrelated with Body Mass Index (BMI) in the elderly with type 2 diabetes so obesity is not the major cause (Table 2).
Figure 17: Frequency distribution in a cohort (n=204), split up to gender of the Netherlands Brain Bank of morbidity of “brain diseases” as ultimate biomarkers of aging further split up to age (Courtesy: Evert de Vries).
Table 2: List of the 19 biomarkers used to define signatures.
As stated earlier, it is estimated that Human lifespan variation is mainly determined by environmental factors, whereas the genetic contribution is 25–30% and expected to be polygenic [2]. The process of aging may be inevitable; however, one many potentially delay the effects and severity of this progression of cognitive decline by focus on these 70%-75% environmental factors which are in common for a healthy life and lay in four domains:
i. Mental exercise: high level of education & stay intellectually engaged;
ii. Physical exercise: avoid a sedentary lifestyle;
iii. Social environment: maintain a social and friendship network;
iv. Healthy diet: including omega-3 fatty acids and protective antioxidants [77].
A Systems Biology approach (Metabolomics: mainly genomics proteomics and lipidomics for brain) can in future lead to a “Personalized medical treatment” and biomarkers of aging. Neuroimaging-biomarkers of Mild Cognitive Impairment (MCI) allow an early diagnosis in preclinical stages of mild-Alzheimer’s disease (m-AD) and dementia. With an aging population in the developed countries emphasis on neuroimaging and smart ligand Pet-scan biomarkers should take place. A second topic of interest –which is still a hypothesis- is the nearly unexploited research area of cognitive reserve. The ability of an individual to demonstrate no cognitive signs of aging despite an aging brain is called cognitive reserve [78,79]. This hypothesis suggests that two patients might have the same brain pathology, with one person experiencing noticeable clinical symptoms, while the other continues to function relatively normally. Studies of cognitive reserve exploring the specific biological, genetic and environmental differences which make one person susceptible to cognitive decline, and allowing another to age more gracefully and in studying this topic biomarkers of aging are clearly warranted.
Albert-Frans Welmers and Evert de Vries, Bluegreentechnologies, The Netherlands for support. Dr. M. Kooreman of the Netherlands Brain bank, Institute for Neuroscience, Amsterdam, The Netherlands is kindly acknowledged for selection of data.

























