info@biomedres.us
+1 (502) 904-2126
One Westbrook Corporate Center, Suite 300, Westchester, IL 60154, USA
Site Map
Received: June 08, 2017; Published: June 16, 2017
Corresponding author: Richard W Cole, New York State Department of Health, Wadsworth Center & Department of Biomedical Sciences; School of Public Health; State University of New York at Albany, Albany, New York, USA
DOI: 10.26717/BJSTR.2017.01.000140
3D cell culture has great potential in the field of tissue engineering and regenerative medicine. Three- dimensional ECM-like substances such as Matrigel® mimic natural proteins found in the ECM and result in a more accurate reflection of cell growth within the human body. In our lab, we compared HT1080 cells transfected with the histone H2B-green fluorescent protein (GFP) growing directly on glass (2D) to those in Matrigel® on the basis of cell growth, proliferation, division and morphology. During cell counting, the cells were maintained at a near-homeostatic temperature, humidity and CO2 level in a custom stage top environmental chamber [1] while being imaged on an Olympus (Melville, NY) IX70 inverted fluorescence microscope. Our results indicated that cell growth, division and proliferation were of greater magnitude in Matrigel® than on a glass coverslip (control). For ultrastructure imaging purposes, a culture of MDCK cells (Canis familiaris, kidney, normal) that were transfected with a pEYFP-Tubulin plasmid subcellular localization vector were used.
Widefield epifluorescent analysis indicated that there was no significant difference in cellular ultrastructure on the basis of flat/rounded morphology and microtubule distribution between the cells grown in glass-bottomed Petri dishes and those in Matrigel®. These results are consistent with our hypothesis that cells in Matrigel® would exhibit greater cell growth, proliferation, and division while maintaining normal cell morphology.
Keywords: Imaging; Fluorescence; Tissue-culture
The importance of culturing in three-dimensional extracellular matrix substitutes has become increasingly evident over the past few years. It stems from that fact it is unnatural for a single cell type to grow in a monolayer on a glass-bottomed Petri dish (MatTek Inc., Ashland, MA). Cells typically grow in multiple layers to form tissues for organs in organ systems that make up complex organisms. These layers often contain a heterogeneous mixture of cells that secrete various chemicals and interact with other cells in their shared environment. Thus, 3D matrices that support multilayer systems are valuable tools for the following: furthering the understanding of cell biology [2]; satisfying the need for cellular models that mimic the functions of living tissues [3], natural conditions for cell response [4] and in vivo environments for 3D cell culture applications. These applications have been explored in a number of ways, including (but not limited to): cell invasion [5] and toxicity assays [6]; spheroid [7], colonoid [8] and stem cell culture [9]; tissue formation [10], engineering [11] and regeneration [12] for therapeutics [13]; and organ development [14].
As the benefits of 3D culture become more widely accepted and utilized, many different ECM substitutes are being developed with the potential to mimic cell behavior as it occurs in vivo. From everyday egg whites to commercial compounds such as CyGEL® (BioStatus, Leicestershire, UK), Matrigel® (Corning Inc., Corning, NY), and a plethora of other hydrogels, the variations of threedimensional matrices are endless. Our experiments led us to discover insurmountable issues with highly variable egg whites and chemically unfixable CyGEL® (personal communication). Matrigel® was ultimately chosen because of its fixable nature, allowing proteins and macromolecules to be labeled. Such cellular components were viewed in order to test the effectiveness and viability of the various gelatinous compounds in mimicking natural cell growth conditions on the basis of their cell counts and ultrastructure as compared to those of 2D cultures.
Though it has been noted that 3D culture may mimic the natural conditions for cell growth, there is not much evidence as to whether it is more effective than 2D culture in this regard. Simply providing space for cells to stratify does not ensure that they will grow at a faster rate, have a higher peak number of cells, or exhibit any changes in morphology. However, changes in proliferation and ultrastructure are to be expected as the cells are given a greater volume in which to grow above and below each other. Thus, the differences between 2D and 3D culture were quantified by the following criteria:
a) Peak number of cells,
b) Rate of cell growth and proliferation, and
c) Cellular ultrastructure.
To our knowledge, no such differences have yet been quantified in this manner.
Matrigel®, a thermo reversible protein-based hydrogel, was aliquoted it into 1.5 mL sterile microcentrifuge tubes and stored in a freezer at -20˚C until use. As Matrigel® is a liquid at temperatures below 22˚C, it was kept in a small bucket of ice in the hood to allow for pipetting. 160 μL of Matrigel® was pipetted into a “silo” (created as shown in Figure 1A), allowing the gelatinous material to assume the shape of the silo. The silo device (see Figure 1B) consists of the top 1 cm of a 1.5 μL microcentrifuge tube that is placed in the center of a 35 mm glass-bottomed Petri dish (MatTek, Inc., Ashland, MA). The device was placed on an aluminum block to increase thermal conductivity between the ice and the silo, thereby preventing the Matrigel® from coagulating. In addition, the silo was kept dry and sterile as the aluminum separated it from the wet ice. (Figure 1A Silo cut from microcentrifuge tube. B) Image of two silos on top of aluminum block inside ice bucket.
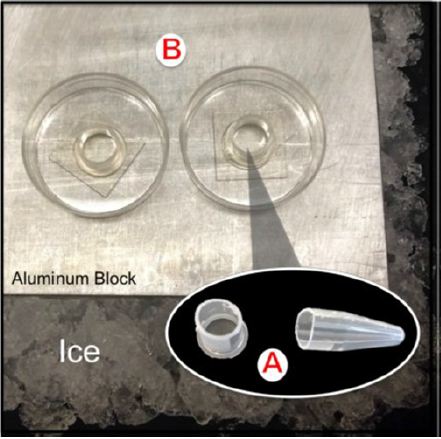
Figure 1: A) Silo cut from microcentrifuge tube. B) Image of two silos on top of aluminum block inside ice bucket.
Basic cell culture and sterile technique were executed using HT1080 cells (Homo sapiens, connective tissue, fibro sarcoma) transfected with the histone H2B-green fluorescent protein (GFP). After trypsinization, 10 mL of phenol red-free DMEM Media® (Invitrogen, Carlsbad, CA) were added to the flask. The mixture of media and cells was pipetted into a 15 mL conical centrifuge tube for centrifugation at 400 g (rcf) for 5 min. After forming a pellet, the cells were resuspended in 1-2 mL of media (depending on the size of the pellet). 10-15 μL of cells were pipetted into each of the two silos, one with Matrigel® and the other without as the control. 160 μL of media were added to the silo (Figure 1A) without Matrigel® and both silos were placed inside a 100 mm Petri dish to allow for gas exchange in the incubator. After the Matrigel® semi-solidified in the incubator for 30-60 min at 37˚C with 5% humidity, 160 μL of media was pipetted into the Matrigel® silo as well. The cells were refed with the phenol-free media every other day.
For imaging purposes, a culture of MDCK cells (Canis familiaris, kidney, normal) that were transfected with a pEYFP-Tubulin plasmid sub cellular localization vector was used to remove the issue of antibody penetration through the Matrigel®. The above procedure was followed with the exception of centrifugation in order to increase viability. After trypsinization with 1 mL of ATV, 16 μL of cells were instead pipetted into each of the two silos, one with Matrigel® (experimental) and the other without (control).
A comparison was made between the HT1080 cells growing directly on glass (2D) and in Matrigel® (3D). The cells were maintained at a near-homeostatic temperature, humidity and CO2 level in a custom stage top environmental chamber [1] while being imaged on an Olympus (Melville, NY) IX70 inverted fluorescence microscope using a 10X (0.30 NA) objective lens and GFP/FITC filter cube. The microscope and camera were controlled with ImagePro® (Medi Cybernetics, Silver Spring MD). The monolayer control was imaged in a single 2D plane whereas the 3D Matrigel® images were acquired as Z-series (200-400 μm depth, 10μm spacing) daily, until the cells became too dense to be counted. When there was no longer definition between nuclei, it became impossible to distinguish individual cells. Thus, our data collection stopped at Day 9. Cell counts were subsequently obtained using the 3D cell counter program in Fiji [15], an image processing application. The sequence of images from the Z-series was projected in 2D on the basis of maximum intensity
The following differences in ultrastructure between the MDCK cells growing in the Matrigel® and those of the control were examined: (1) flat/rounded cellular morphology and (2) microtubule distribution. The primary variation in the indirect immunofluorescence procedure was in the duration of Hoechst 33342 staining: 5 min for the control vs. 10 min to accommodate Matrigel® penetration. The cells of the control were rinsed three times for three minutes each in phenol red-free DMEM Media® at 37˚C with 5% humidity. The cells growing in Matrigel® were rinsed three times for five minutes each to accommodate the slower penetration rate. The analysis of ultrastructure was done using a Nikon TE2000 Inverted Microscope.
The mean ratio (N=5) of 2D cell counts in a Matrigel® monolayer to those on a glass-bottomed Petri dish was calculated over a period of a few days, resulting in a ratio of about 1. Thus, it was empirically determined that the differences in cell growth and proliferation were due to the 3D property of Matrigel® as opposed to its chemical composition. To compare changes between 2D and 3D proliferation, the mean ratio (N=5) of Matrigel® cell counts to control cell counts was calculated for each day. As Figure 2 demonstrates, the ratio consistently exceeds 1 by a value greater than 0.5.
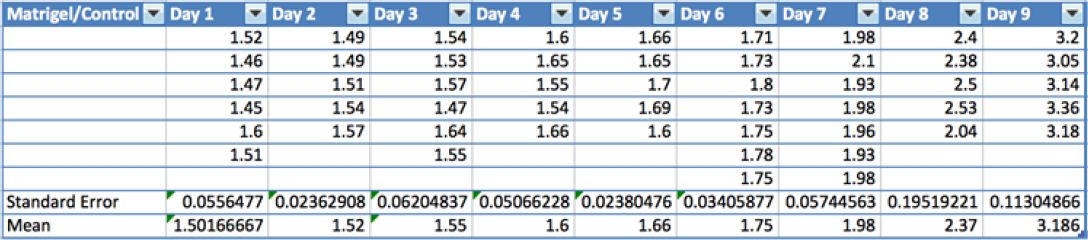
Figure 2: Table of normalized HT1080 cell counts in 3D Matrigel vs. 2D control over 9-day period. Each table Column reflects 5-9 samples with the mean and standard error of each set denoted below for each day.
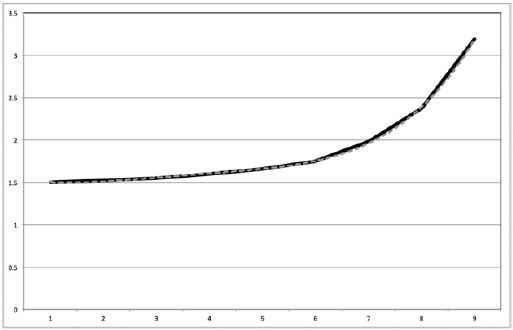
Figure 3: Exponential trend line (dotted) for normalized counts of HT1080 cells in Matrigel® vs. control over 9-day period (solid).
The positive slope of the curve in Figure 3 confirms that the cell counts in 3D Matrigel® were consistently higher than those of the 2D control. Furthermore, the upward curvature (rapidly increasing slope) indicates that a greater number of cells in Matrigel® were dividing compared to the control. The cells of the control reached their peak number after 6 days on average whereas the cells in Matrigel® had a higher carrying capacity, lasting over 9 days. This claim is corroborated by the upward spike in the ratio shown by Figure 3 after 6 days.
As shown in Figure 4, there was no significant difference in cellular ultrastructure based on flat/rounded morphology and microtubule distribution between the cells grown in glass-bottomed Petri dish and those in Matrigel®. The cells appeared viable as they were in varying states of mitosis. One of the control cells (located left-middle) was undergoing prophase with two spindle poles clearly visible. Similarly, one of the cells in Matrigel® (bottommiddle) appeared to be in late anaphase. Though some of the cells appeared to lack microtubules, this phenomenon was likely a result of the transgenic cells’ inconsistency in the expression of GFP.
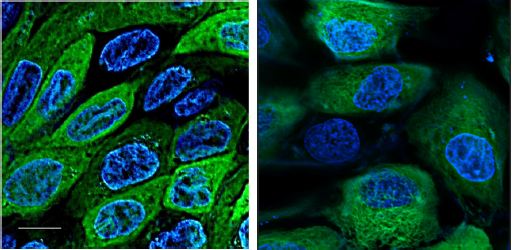
Figure 4: A slice of a Z-series in the control (left) and experimental (right). MCDK cells imaged in EPIfluorescent mode, labeled for microtubules (green) and DNA (blue). Bar = 10 μm.
Though inexpensive and easily obtainable, egg whites were found to be highly variable in our laboratory. The amount of time required heating the egg white to an ideal semi-solid state ranged from 30-90 min and the appropriate viscosity was difficult to identify. After many trials, we determined that the results from any single 3D egg white culture are largely irreproducible. The auto fluorescence of the egg white also leads to problems with imaging. To our knowledge, there is only one paper [16] that cites this method (Kaipparettu et al.), further indicating its inadequacy for this application.
Conversely, Matrigel® yielded reproducible results. This predictability is largely due to the fact that Matrigel® consistently hardens after being incubated at 37˚C with 5% humidity for 30-60 min. Moreover, Matrigel® is chemically fixable, allowing for protein tagging and fluorescent imaging of the cytoskeleton. CyGEL®, though similarly thermo-reversible, is unfixable. It is not only extremely miscible in H2O-based solutions such as media, but also lacks the ability to be fixed. Matrigel® was clearly the most suitable ECM substitute.
The voluminous Matrigel® provides a large space for cell growth as opposed to the flat plane of the glass cover slip (control). The ability to grow out in three dimensions allows for a greater number of cell divisions per unit area in Matrigel® as opposed to the glassbottom of the control dish. Matrigel® allows for a greater surface area of cell-to-cell contact without limiting nutrient absorption. Supported in all directions, the cells grow around each other with enough of the ECM between them for sustenance. In addition, the cushioning effect of the ECM limits the effect of convergent physical forces. On the contrary, the cells of the control have no such inhibition. They overgrow into a compact mass, suffocate and die from lack of nutrients. The malignant mound contains indistinct irregular cells. Healthy cells are naturally more rounded [17].
Matrigel® maintains euplasia – normal cell morphology. A comparison between the microtubules and DNA of the control cells to those of the cells grown in Matrigel® showed them to be virtually indistinguishable. This outcome is not surprising given the aforementioned physical properties and capabilities of Matrigel® that make it conducive to cell growth. The future of 3D cultures lies in regenerative medicine. It may be used to build tissues, or even generate organs. Natural hydrogels such as protein-based Matrigel® are ideal scaffolds for tissue engineering due to their biocompatibility and inherent biodegradability [18]. Our results corroborate the viability of Matrigel® as an ECM substitute in vitro on the basis of its physical properties as well as its positive effects on cell growth, proliferation and morphology. In addition, hydrogels may be ideal for transplantation of cells due to their low toxicity and high diffusivity of biomolecules at body temperature [19]. However, since Matrigel® was isolated from the basement membrane of a mouse sarcoma, it is unlikely to be used in clinical trials. Other hydrogels or matrices may need to be used depending on the cell type and end product desired.
This work was supported by the New York State Department of Health, Wadsworth Center’s Advanced Light Microscopy & Image Analysis Core Facility.

























