 Review Article
Review ArticleAbstract
In order to study the role of various nuclei of the hypothalamus in the implementation of the drinking skill developed electrostimulation and temporary shutdown were performed supraoptic (SO), ventromedial (VMH) and medial-mamillary (MM) nuclei of the hypothalamus. The registration of the amount of consumed water showed the following: electrostimulation of SO, unlike VMH and MM of hypothalamic nuclei caused additional water consumption in comparison with the background. The application of novocaine in SO led to a significant decrease in the amount of water consumed, which decreased for the duration of the action of novocaine (40-50 minutes) and amounted to 60-70 ml instead of 120 in the norm. Bilateral novocaine blockade of SO led to a complete inhibition of the performance of the developed drinking skill. Thus, the results of the studies carried out on electrical stimulation and temporary switching off of the SO suggest that the osmoreceptors of the SO of the anterior hypothalamus play a certain role in the processes of the formation of thirst.
Keywords: Various Nuclei of the Hypothalamus; Drinking Conditioned Reflex; Electrical Stimulation; Temporary Shutdown
Abbreviations: SO: Supraoptic Nucleus of the Hypothalamus; VMH: Ventromedial Nucleus of the Hypothalamus; MM: Medial-Mammillary Nucleus of the Hypothalamus
Introduction
The hypothalamus is a small area in the diencephalon that includes a large number of cell groups (more than 30 nuclei) that regulate the neuroendocrine activity of the brain and homeostasis of the body [1-4]. The hypothalamus is connected by nerve pathways with almost all parts of the central nervous system, including the cortex, hippocampus, amygdala, cerebellum, brain stem, and spinal cord [5]. Together with the pituitary gland, the hypothalamus forms the hypothalamic-pituitary system, in which the hypothalamus controls the synthesis and secretion of many pituitary hormones, is the central link between the nervous and endocrine systems. It secretes hormones and neuropeptides and regulates functions such as hunger and thirst. body thermoregulation, sexual behavior, sleep and wakefulness (circadian rhythms) [1,2,6].
Recent studies show that the hypothalamus plays an important role in higher functions, such as memory and emotional state, and thus participates in the formation of various aspects of behavior [2,7,8]. The hypothalamus controls the activity of the human endocrine system, due to the fact that its neurons are able to secrete neuroendocrine transmitters (liberins and statins) that stimulate or inhibit the production of hormones from the adenohypophysis [3,5]. Suppression of the functional activity of the hypothalamus leads to a violation of drinking and eating behavior [9]. Considering all of the above, the task of this study was to study the role of various nuclei of the hypothalamus in the implementation of the drinking skill developed.
Methods
Experiments were performed using 20 rabbits breeds of “Chinchilla” in weight 2-3kg trained to perform a conditioned operant drinking behavior reflex. The animals were subjected to water deprivation for 24 and 48 hours, after which they were trained in the instrumental drinking skill. In response to the sound signal, the rabbit pressed the pedal, as a result of which the door opened, jumped over the barrier from the starting compartment of the chamber to its target section to receive water in a strictly defined dose (5-10 ml) and then returned back to the starting section cameras. The conditioned stimulus was applied at regular intervals (every 45 s) from 10 to 15 times during the experiment. During the experimental day, the animals received an average of 100-120 ml of water. The experiments were carried out under conditions of 100% reinforcement. In the study of behavioral reactions, the time from the moment the signal was given to the start of the jump (latency period), the time of running, lapping, and also the time spent on returning to the starting box of the camera were recorded.
Electrical stimulation of the supraoptic - SO (A-3; L2.2; H15.8), ventromedial - VMH (P1; L0.5; H17) and medial-mammillary - MM (A-3; L0.5; H18.5) nuclei of the hypothalamus of the brain was carried out using a universal electrostimulator ESU-1.
Results and Discussions
Observations of conditioned reflex activity before the start of the experiment showed that the time spent on the execution of individual links of the developed drinking habit, on average, was: the latent period of reaction to the conditioned stimulus - 1.2±0.07 sec, the time of jumping and running – 3.2 ± 0.07 sec, drinking time - 19.8±0.17 sec, return time to the starting compartment of the chamber was 3.97±0.09 sec. Electrical stimulation of SO, VMH and MM nuclei of the hypothalamus at low parameters of stimulation (60-100 μA, 5 Hz, 0.5 ms) did not effect on the performance of the drinking skill. An increase in stimulation parameters (100-300 μA, 5-100 Hz, 0.5 ms) also did not lead to significant changes in the behavior of animals, with the exception of the latent period of the conditioned reflex, which increased from 1.2 ± 0.07 to 3.17±0.06 sec. The time spent on running jump, lap up and return remained unchanged and averaged 3.2± 0.07, respectively; 19.7±0.15; 3.9±0.08 sec. The amount of water consumed in the experiment did not change and remained at the background level (100-120 ml).
The conditions of the conditioned reflex described above are presented by the example of stimulation of the VMH of the hypothalamic nucleus. Similar changes in the behavior of animals were observed upon electrical stimulation of the nuclear MM. A somewhat different behavior of rabbits was observed upon stimulation of the SO nucleus of the hypothalamic. Electrical stimulation of the SO nucleus of the hypothalamic caused the appearance of signs of emotional anxiety. In connection with licking and sniffing the starting section of the chamber, the latent period of the reaction slightly increased from 1.2 ± 0.07 to 3.23 ± 0.07 sec. The time spent on jumping and running, water consumption and return remained unchanged, which averaged 3.3 ± 0.07, respectively; 20.07 ± 0.15; 4.13 ± 0.09 sec.
The registration of the amount of consumed water showed the following: electrostimulation of SO caused additional water consumption in comparison with the background. The amount of water drunk during the entire period of the experiment doubled and amounted to 200-250 ml (Figure 1).
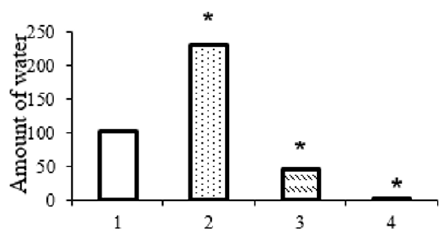
Figure 1: Change in the amount of water consumption as a result of electrostimulation and novocaine blockade of the supraoptic nucleus of the hypothalamus. 1 - background, 2 - after electrical stimulation, 3.4 - after unilateral and bilateral novocaine blockade of the supraoptic nucleus of the hypothalamus, respectively.
Stimulation of the VMH and MM of the hypothalamic nuclei did not interfere with the performance of the developed drinking habit in animals previously deprived of water, the latent period of the reaction to the conditioned stimulus slightly increased. The amount of water consumed did not change in comparison with the background. The introduction of a 5% solution of novocaine into the VMH and MM of the hypothalamic nucleus did not interfere with the performance of the developed drinking habit. At the same time, the temporal parameters of the skill performance remained at the level of the background indices and were for the VMH: the latent period of reaction to the conditioned stimulus was on average 1.2 ± 0.07 sec, the jump and run from the starting section of the chamber to the target - 3, 27 ± 0.07 sec, drinking time - 20.13 ± 0.09 sec and return to the starting compartment - 4.23 ± 0.7 sec. Similar data observed in the behavior of animals were obtained after the introduction of a 5% solution of novocaine into the MM of the hypothalamic nucleus.
A somewhat different behavioral picture was observed with the introduction of 5% solution of novocaine into the SO of the anterior hypothalamus. The animal reacted to the sound signal with a long latency period: 3.97,90.11 instead of 1.27 ± 0.07 sec. The time spent on running jump also increased - from 3.23 ± 0.07 to 5.5 ± 0.08 sec, water consumption - from 20.07 ± 0.17 to 32.5 ± 0.33 sec and return - from 4.1 ± 0.1 to 7.53 ± 0.1 sec. The application of novocaine in SO led to a significant decrease in the amount of water consumed, which decreased for the duration of the action of novocaine (40-50 minutes) and amounted to 60-70 ml instead of 120 in the norm. Bilateral novocaine blockade of SO led to a complete inhibition of the performance of the developed drinking skill (Figure 1).
Observation of the behavior of animals revealed the following. Electrical stimulation of the VMH and MM of the hypothalamus did not cause disturbances in the behavior of the animals. Only the latency period of the reaction increased slightly. With electrical stimulation of the nucleus SO, the animals showed signs of emotional anxiety, and when a sound signal was applied, the developed drinking habit was retained, but the amount of water drunk significantly increased (2 times). Anderson [10] observed a similar increase in the drinking reaction upon irritation of the anterior nuclei of the hypothalamus in contrast to the data of other authors, who noted an increase in water consumption during electrical stimulation of the lateral hypothalamus [11], irritation of the preoptic region [12], as well as destruction of the VMH of the hypothalamic nucleus in rats [13].
In our experiments, various effects of novocaine blockade of the studied nuclei of the hypothalamus were revealed. So, if the blockade of VMH and MM did not cause an effect on the developed drinking habit, then even one-sided switching off of the SO of the nucleus caused an increase in all time parameters. The amount of water consumed has significantly decreased. Bilateral application of novocaine to this nucleus led to a complete inhibition of the performance of the developed drinking habit and refusal of water, despite the fact that before coagulation the animals had a pronounced drinking motivation. Complete adipsia and a sharp decrease in water consumption after the destruction of the anterior nuclei of the hypothalamus were observed by a number of authors [10], in the opposite of the data of other researchers who caused adipsia when the lateral hypothalamus was damaged [14], with the joint destruction of the lateral hypothalamus and the lateral preoptic region [15], as well as with damage to the medial hypothalamus. The appearance of thirst upon electrical stimulation of the SO of the anterior hypothalamus and its disappearance with bilateral novocaine blockade of this area, apparently, can be explained by the presence of osmosensitive neurons in the SO, which were found in this nucleus [16] and to which some researchers assign a primary role in the regulation of water consumption [17].
Thus, the results of the studies carried out on electrical stimulation and temporary switching off the SO suggest that the osmoreceptors of the SO of the anterior hypothalamus play a certain role in the processes of the formation of thirst.
References
- Sokol NV (2014) Triticale is a grain culture. Palmarium Academic Pulishing pp. 143.
- Pankratov GN, Meleshkina EP, Kandrokov RKh, Vitol IS (2016) Technological properties of new varieties of triticale flour. Khleboprodukty 1: 60-62.
- Vitol IS, Karpilenko GP, Kandrokov RKh, Starichenkov AA, Koval AI, et al. (2015) Protein-proteinase complex of triticale grain. Storage and processing of agricultural raw materials 8: 36-38.
- Vitol IS, Meleshkina EP, Kandrokov RKh, Verezhnikova IA, Karpilenko GP (2016) Biochemical characteristics of new varieties of triticale flour. Khleboprodukty 2: 42-43.
- Magomedov GO, Malyutina TN, Shapkarina AI (2016) Development of technology for whipped flour confectionery products of increased nutritional value using triticale flour. Vestnik VSUIT 1: 106-109.
- Karchevskaya OV, Dremucheva GF, Grabovets AI (2013) Scientific foundations and technological aspects of the use of triticale grain in the production of bakery products. Bread-making of Russia 5: 28-29.
- Woś H, Brzeiński W, Woś J (2013) Breadmaking quality Triticale bred in Poland. 8th International Triticale Symposium, Ghent, Belgium pp. 23.
- Koryachkina SYa, Kuznetsova EA, Cherepnina LV (2011) Technology of whole grain triticale bread. Orel: GU-UNPK pp. 116.
- Grabovets AI, Krokhmal AV, Dremucheva GF, Karchevskaya OE (2013) Breeding of triticale for baking purposes. Russ. Agric. Sci 39: 197-202.
- Rakha A, Aman P, Andersson R (2013) Rheological characterization of aqueous extracts of triti-cale grains and its relation to dietary fiber characteristics. J Cereal Sci 57(2): 230-236.
- Rakha A, Aman P, Andersson R (2013) Rheological characterization of aqueous extracts of triticale grains and its relation to dietary fiber characteristics. J. Cereal Sci 57: 230-236.
- Meleshkina EP, Pankratov GN, Vitol IS, Kandrokov RH, Tulyakov DG (2017) Innovative Trends in the Development of Advanced Triticale Grain Processing Technology. Foods and Raw Materials 5(2): 70-82.
- Pankratov GN, Meleshkina EP, Vitol IS, Kandrokov RKh (2017) Actual directions of technological development of the milling branch of the food industry. Food industry 8: 44-49.
- Barnett RD, Blount AR, Pfahler PL, Bruckner PL, Wesenberg DM, et al. (2006) Envi-ronmental stability and heritability estimates for grain yield and test weight in triticale. J Appl Genet 47: 207-213.
- Blum A (2014) The abiotic stress response and adaptation of triticale - a review. Cereal Res. Com-mun 42(3): 359-375.
- De Laethauwer S, Reheul D, De Riek J, Haesaert G (2012) Vp1 expression profiles during kernel de-velopment in six genotypes of wheat, triticale and rye. Euphytica 188: 61-70.
- Dennett AL, Cooper KV, Trethowan RM (2013) The genotypic and phenotypic interaction of wheat and rye storage proteins in primary triticale. Euphytica 194: 235-242.
- Manley M, McGoverin C, Snyders F, Muller N, Botes W, et al. (2013) Prediction of triticale grain quality properties, based on both chemical and indirectly measured reference methods using near-infrared spectroscopy. Cereal Chem 90(6): 540-545.
- McGoverin C, Snyders F, Muller N, Botes W, Fox G, et al. (2011) A review of triticale uses and the effect of growth environment on grain quality. J. Sci. FoodAgric 91(7): 1155-1165.
- He ML, McAllister TA, Hernandez-Calva LM, Aalhus JL, Dugan MER, et al. (2014) Effect of dietary inclusion of triticale dried distillers’ grain and oilseeds on quality and fatty acid profile of meat from feedlot steers. Meat Sci 97(1): 76-82.
- Ukalska J, Kociuba W (2013) Phenotypical diversity of winter triticale genotypes collected in the Polish gene bank between 1982 and 2008 with regard to major quantitative traits. Field-CropsRes 149: 203-212.
- Bona L, Acs E, Lantos C, Purnhauser L, Lango B, Tomoskozi S (2013) Human utilization of tritica-le: technological and features, milling and baking experiments. In: Abstracts 8th international trit-icale symposium. Ghent, Belgium pp. 46.
- (2009) Guide to Mixolab Applications. Rheological and enzymatic analysis (Ma-nueld'applications Mixolab) 28: 79.
- (2008) ICC No. 173. Whole meal and flour from T. aestivum - Determination of rheological behavior as a function of mixing and temperature increase, ICC, Standard. Vienna.
- Antanas S, Alexa E, Negrea M, Guran A, Lazureanu E (2013) Studies regarding rheological proper-ties of triticale, wheat and rye flours. J. of Horticulture, Forestry and Biotechnology 17(1): 345-349.
- Tulyakov DG, Meleshkina EP, Vitol IS, Pankratov GN, Kandrokov RKh (2017) Evaluation of the properties of flour from triticale grain using the Mixolab system. Storage and processing of agricultural raw materials 1: 20-23.
- Dubat A Le mixolab (2009) Profiler: un outilcomplet pour le controlequalite des bles et des farines. In-dustries des Cereales 161: 11-26.
- Dubtsova GN, Nechaev AP, Molchanov MI (2000) Molecular biological aspects of the formation of lipid-protein complexes and assessment of their role in the structure of gluten. In the book: Vegetable protein: new perspectives. Moscow: Pishchepromizdat pp. 100-121.
