Research ArticleOpen Access
Heat Stress Induced Alterations in Prostaglandins, Ionic and Metabolic Contents of Sheep Endometrial Epithelial Cells In Vitro Volume 1 - Issue 4
Received: September 12, 2017; Published: September 22, 2017
Corresponding author: Sukanta Mondal, ICAR-National Institute of Animal Nutrition and Physiology, Adugodi, Bangalore-560030, India
DOI: 10.26717/BJSTR.2017.01.000384
Abstract
Background: Heat stress is one of the major factors responsible for reduced fertility in livestock including sheep. The present study wasaimed to elucidate the effect of heat stress on prostaglandins, ionic and metabolic contents of sheep endometrial epithelial cell in vitro. Sheepwhole genitalia containing ovaries were collected immediately after slaughter and transported to the laboratory. Following washing the healthyand non pregnant uterine lumen with HBSS with gentamycin and 0.1% BSA, HBSS containing 0.3% trypsin was then infused and epithelial cellswere isolated by incubation at 370C for 60min. After cell counting and viability determination, the epithelial cells were cultured in at 38.5oC for24hr (control). Heat stressed cultures were acclimated at 38.50C for 6hr and then placed at 40.50C for 18 hr.
Results: In vitro heat shock significantly (P<0.05) increased protein, phosphorous, urea and PGF2α contents in culture medium whereassignificantly (P<0.05) decreased SOD content as compared to control. Elevated temperature did not significantly (P<0.05) increase calciumcontents but non-significantly (P>0.05) increase glucose, chloride and PGE2 levels in culture medium. It was concluded that heat stress alteredprostaglandins, ionic and metabolic contents of endometrial epithelial cells In vitro
Keywords: Heat stress; Prostaglandins; Endometrial epithelial cells; sheep
Abbreviations: HS: Heat stress; PGs : Prostaglandins; SOD : Superoxide Dismutase Activity
Introduction
Early embryonic mortality is one of the major intriguingfactors of reproductive failure that causes considerable challengeto the mammalian cell biologists. Majority of the losses occurs dueto failure of cellular and molecular dialogues at embryo uterineinterface. Higher environmental temperature is one of the majorfactors responsible for reduced fertility in farm animals [1]. Heatstress (HS) can compromise reproductive events by decreasingthe expression of estrous behavior, altering ovarian folliculardevelopment, compromising oocyte competence, and inhibitingembryonic development. Heat stress also increases the productionof PGF2α in the endometrium, leading to the early regression of CLor the death of embryos [2]. It was observed that heat stress from 8to 16 days after insemination modulated the uterine environment[3], reduced the weight of corpora lutea and impaired conceptusgrowth [4].
Prostaglandins (PGs) produced by endometrium servesas a crucial mediators in maternal recognition of pregnancy,implantation and parturition [5]. The endometrial epithelialand stromal cells have specific morphological and functionalproperties. Epithelial cells preferentially produce PGF2α whereasstromal cells produce mainly PGE2 [6]. PGF2α acts as the luteolyticagent [7] to control the estrous cycle in ruminants. Endometrialsecretion of PGF2a by pregnant uterus has been found to increasein response to heat stress and decrease the embryonic survivalityby altering the signals required for maintenance of corpus luteumfunction during early pregnancy. Increased PGF2a syntheticcapacity of endometrium exposed to heat stress may be due toheat-induced alterations in endometrial cellular membranesresulting in increased mobilization of substrate for prostaglandinbiosynthesis. Similar increases in uterine PGF2a secretion in response to heat stress in vitro by endometrial expiants from cowsat Day 17 of pregnancy have been reported [2]. Furthermore,in-vivo heat stress of gilts between Days 8 and 16 of pregnancyincreased concentrations of PGFM in the peripheral circulationand compromised luteal function, as indicated by reduced plasmaconcentrations of progesterone during Days 13 through 19 [8].The present study was undertaken to investigate the effect of heatstress on prostaglandins, ionic and metabolic contents of sheependometrial epithelial cells In vitro.
Materials and Methods
Isolation of endometrial epithelial cells
Sheep uteri were collected from the local abattoir immediatelyafter slaughter and transported to the laboratory on ice. Theepithelial cells from the sheep endometrium were separated by themethod [9] with slight modification. Briefly, the uterine lumen wasinitially washed with HBSS supplemented with gentamycin and0.1% BSA. Sterile HBSS containing 0.3% trypsin was then infusedinto the uterine lumen. Epithelial cells were isolated by incubationat 370C for 60min. The cell suspension obtained from the digestionwas filtered to remove un-dissociated tissue fragments. The filtratewas washed with HBBS supplemented with gentamycin and 0.1%BSA by centrifugation at 600xg for 10min. The number of viable cellsthat excluded Try pan blue was counted using a hemocytometer.After cell counting and viability determination, the epithelial cellswere seeded at the rate of 1x105 viable cells in RPMI 1640 mediumat 38.5oC in presence of 5%CO2 for 24hr. The viability of epithelialcells at the time of plating was greater than 90%. The medium waschanged every alternate day until the confluence was reached
Exposure of Heat Stress to Epithelial Cells in vitro
Following isolation of epithelial cells, the cells were culturedin RPMI 1640 medium at 38.5oC in presence of 5%CO2. Controlcultures were maintained at 38.50C for 24hr. Control cultures wereincubated under conditions representing normal body temperatureof normal sheep in a thermo neutral environment. Heat stressedcultures were acclimated at 38.50C for 6hr and then placed at40.50C for 18hr. The culture medium was collected after 24hr andstored at 700C until analysis. The experiment was replicated 6times on different days.
Biochemical analysis
The concentrations of metabolites (Glucose, Total Proteinand Urea) and ions (Calcium, Chloride and Phosphorus) wereanalyzed by using Biochrom Libra S32 UV/Vis Spectrophotometer.Commercial kits used for estimations except SOD were from SpanDiagnostics Ltd, Surat, India. Superoxide dismutase activity (SOD)of the samples was measured spectrophotometrically using Ransodkit (Randox Laboratories Ltd., UK). All measurements were carriedout according to the manufacturer’s instructions. The intra andinter assay coefficients of variation for all analyses were below 7%.
Determination of PGF2a and PGE2
The concentrations of PGF2a and PGE2 were determined in 50μlaliquots of culture medium after 10 fold dilution with extraction buffer using ELISA kits supplied by Neogen, USA. The sensitivity ofthe PGF2a and PGE2 assays were 0.002 and 0.1ng/ml respectively.The intra and inter assay coefficients of variation of PGF2a assaywere less than 19%. The intra and inter assay coefficients ofvariation of PGE2 assay were less than 14%. The cross reactivityof the antisera against 6-keto prostaglandin F1a, 13, 14 dihydro-15keto-prostaglandin F2a, prostaglandin D2 and prostaglandin E2 were3.05%, 0.05%, 0.05% and <0.01%, respectively. The cross reactivityof the antisera against 6-keto prostaglandin E1, 13, 14 dihydro-15keto-prostaglandin F2a and prostaglandin D2 were 0.91%, 0.01%,0.01%, respectively.
Statistical Analysis
The experiment was replicated 6 times on different days.Results are expressed as mean SEM. Concentrations of each factorin control and heat stressed culture medium were analyzed by usingGraph Pad Prism 5 (Graph Pad Software Inc., San Diego, CA, USA).
Results
Figure 1a : Protein content in culture medium followingheat stress in vitro (CTR, Control; HS, Heat Stress).Different superscript letters indicate a significant difference (P<0.05; ANOVA).
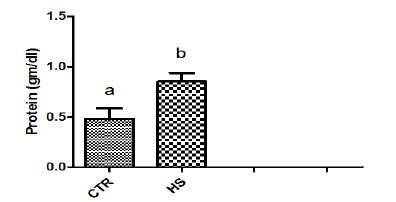
Figure 1b : Phosphorus content in culture mediumfollowing heat stress in vitro (CTR, Control; HS, HeatStress). Different superscript letters indicate a significantdifference (P<0.05; ANOVA).
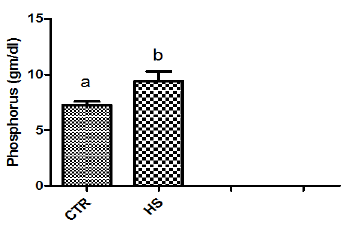
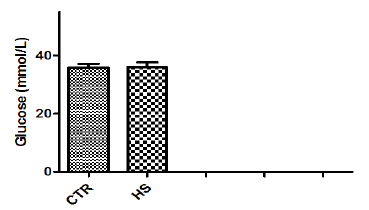
The effect of heat stress on endometrial ionic, metaboliccontents and prostaglandin profiles (Figure 1a-1i). The elevatedtemperature significantly (P<0.05) increased protein, phosphorous,urea and PGF2α contents but significantly (P<0.05) decreased SODcontent in culture medium as compared to control. Exposure ofepithelial cells to 40.50C decreased (P>0.05) calcium but increased(P>0.05) glucose, chloride and PGE2 levels in culture medium.
Figure 1c : PGF2α content in culture medium following heatstress in vitro (CTR, Control; HS, Heat Stress). Differentsuperscript letters indicate a significant difference (P<0.05;ANOVA).
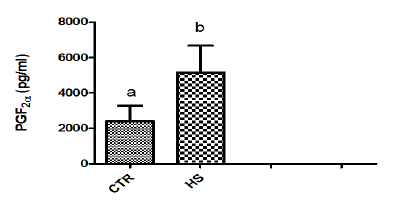
Figure 1d : Urea content in culture medium following heatstress in vitro (CTR, Control; HS, Heat Stress). Differentsuperscript letters indicate a significant difference (P<0.05;ANOVA).
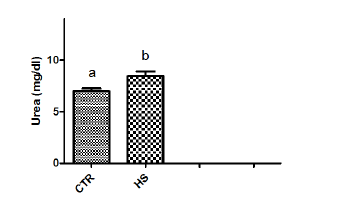
Figure 1e : SOD content in culture medium following heatstress in vitro (CTR, Control; HS, Heat Stress). Differentsuperscript letters indicate a significant difference (P<0.05;ANOVA).
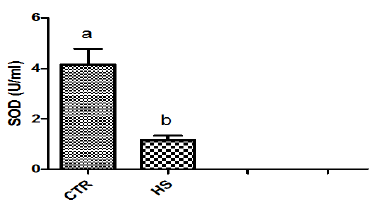
Figure 1f : Calcium content in culture medium followingheat stress in vitro. (CTR, Control; HS, Heat Stress).Different superscript letters indicate a significantdifference (P<0.05; ANOVA).
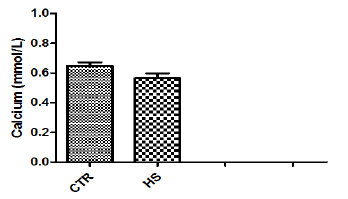
Figure 1g : Chloride content in culture medium followingheat stress in vitro (CTR, Control; HS, Heat Stress).Different superscript letters indicate a significantdifference (P<0.05; ANOVA).
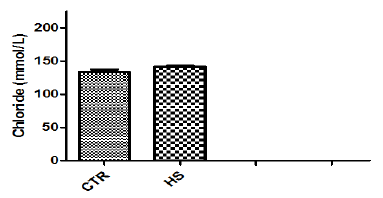
Figure 1h : PGE2 content in culture medium following heatstress in vitro (CTR, Control; HS, Heat Stress). Differentsuperscript letters indicate a significant difference (P<0.05;ANOVA).
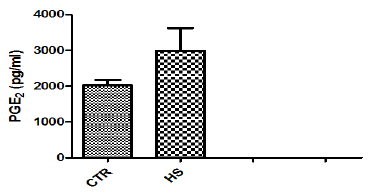
Figure 1i : Glucose content in culture medium followingheat stress in vitro (CTR, Control; HS, Heat Stress).Different superscript letters indicate a significantdifference (P<0.05; ANOVA).
Discussion
Higher environmental temperature is one of the major intriguingfactors responsible for infertility in farm animals. Suppressionof reproductive function by heat stress is marked by abnormalendometrial function [10], compromised follicular growth [11],alteration in hormonal secretion [12] and embryonic function[13]. The present study was undertaken to delineate the effect ofin vitro heat stress on prostaglandin production in sheep. In vitroheat stress mediated alterations in ionic and metabolic contentsof endometrial epithelial cells were also analyzed. In the present,in vitro elevation of incubation temperature of sheep endometrialepithelial cells from 38.50C to 40.50C increased the productionof protein. Our results agree with earlier reports [2] in terms of increase in protein secretion by conceptus and endometriumduring early pregnancy in bovines. This may be due to enhancedability of endometrium to regulate the rate of metabolic activity at40.50C. In vivo thermal stress has been found to enhance in vitroprotein production by conceptus in pig [14].
This may be due to fact that high environmental temperaturemay severely alter metabolic activity and lead to failure of conceptusto produce biochemical signals in adequate amount required forpreventing CL regression. Heat shock results in specific changes inthe patterns of protein synthesis by mammalian cells, characterizedby the synthesis of a small number of intracellular proteins referredto as heat-shock or stress proteins that may provide a degree oftolerance to stress [15]. In the current study, in vitro heat stressincreased phosphorous and urea contents in culture medium. Ourresults are in agreement with the report of Daniel and Korsmeyer[16] who suggested that any stress conditions induces the influx ofions and metabolites particularly calcium, magnesium, phosphorus,chloride as well as glucose and glucose regulated proteins [17].Significant increase in PGF2α contents was observed in epithelialcells exposed to 40.50C as compared to those incubated at 38.50C(control). Our results agree with the report of Putney [2] andMalayer [10] wherein they suggested that secretion of both PGF2αand PGE2 in endometrium increased in response to heat stress.
Exposure of heat shock in the present study resulted inmarked increase in the release of PGF2α into culture medium dueto alterations in membranes resulting in increased mobilization ofsubstrates for prostaglandin biosynthesis. Heat induced increasein the turnover of membrane phospholipids and the releaseof arachidonic acid may provide substrates for prostaglandinsynthesis [18]. As maintenance of luteal function is associated withalterations in endometrial prostaglandin production [19], increasedprostaglandin secretion following heat stress may compromiseCL function and initiate luteal regression. Elevated temperaturedecreased the activity of the endometrial inhibitor of PG synthesispresent at Day 17 of pregnancy. Putney [20] found similar resultswhen the endometrium was subjected to elevated temperature andthe isolated inhibitor was tested for activity. Reduced function ofthis proteinaceous inhibitor of the cyclooxygenase-endoperoxidaseenzyme complex might account partially for increased PGF2αsecretion in pregnant animals but cannot account for heat-inducedPGF2α release from endometrium of cyclic animals.
The temperature induced increase of PGF2α secretion by theendometrium was also not due to a direct effect of temperature onreaction rate of cyclooxygenase activity, because the productionof PGF2α from a cell-free preparation of cyclooxygenaseendoperoxidaseenzyme complex from periparturient cotyledonwas depressed by elevated temperature. In the present study, thelevels of PGE2 did not significantly (P>0.05) increase in epithelialcells exposed to 40.50C as compared to control group. Our resultsare in agreement with the reports of [2,20] wherein elevatedtemperature did not increase PGE2 secretion, indicating thatelevated temperature affects PG secretion in some manner specificfor PGF2α. At Day 17, most PGF2α is released from the endometrial epithelium, while most of the PGE, and originates in the stroma [6].Perhaps high temperature affects these two cell types differently orpreferentially enhances the activity of endoperoxide F reductase.Similarly, Kobayashi [21] suggested that elevated temperaturedid not affect PGE2 production in isthmic epithelial cells of cattleoviduct.
They suggested that the synthesis of PGE2 was not affectedby elevated temperatures in the isthmus due to the presence ofdifferent cell type populations (ciliated and secretary). Our resultsdiffered from the report of [21] wherein they suggested that PGE2production was increased by elevated temperature in ampullaryepithelial cells of cattle oviduct and the increased PGE2 productioncould be due to either increases in the expressions of PGESs orincreases in the PGES activity. It is concluded that in vitro heatstress altered prostaglandins, ionic and metabolic contents ofsheep endometrial epithelial cells. These in vitro results suggestthat exposure of uterine epithelial cells to high environmentaltemperature may disrupt the endometrial factors responsible formaintenance of pregnancy.
Acknowledgement
The authors are grateful to National Fund for Basic, Strategicand Frontier Application Research in Agriculture (Grant No. NASFAS2002), Ministry of Agriculture, ICAR, New Delhi for providingfinancial support to carry out this work. The authors are thankful toDr. P.K. Agarwal, ADG (NASF) and Dr. A. Bandyopadhyay, Ex NationalCoordinator, NFBSFARA for their help and constructive suggestions.We thank the director, NIANP, for providing the necessary facilitiesto carry out the research work. The help rendered by A. Jagannathis duly acknowledged.
References
- Gwazdauskas FC, Thatcher WW, Wilcox CJ (1973) Physiological, Environmental,and Hormonal Factors at Insemination Which May AffectConception. J Dairy Sci 56(7): 873-877.
- Putney DJ, Malayer JR, Gross TS, Thatcher WW, Hansen PJ, et al. (1988)Heat stress-induced alterations in the synthesis and secretion of proteinsand prostaglandins by cultured bovine conceptuses and uterineendometrium. Biol Reprod 39(3): 717-728.
- Geisert RD, Zavy MT, Biggers BG (1988) Effect of heat stress on conceptusand uterine secretion in the bovine. Theriogenol 29(5): 1075-1082.
- Biggers BG, Geisert RD, Wetteman RP, Buchanan DS (1987) Effect of heatstress on early embryonic development in the beef cow. J Anim Sci 64(5):1512-1518.
- Poyser NL (1995) The control of prostaglandin production by the endometriumin relation to luteolysis and menstruation. Prostaglandins LeukotEssent Fatty Acids 53(3): 147-195.
- Fortier MA, Guilbault LA, Grasso F (1988) Specific properties of epithelialand stromal cells from the endometrium of cows. J Reprod Fertil83(1): 239-248.
- Bazer FW, Ott TL, Spencer TE (1994) Pregnancy recognition in ruminantspigs and horses: signals from the trophoblast. Theriogenol 41:79-94.
- Hoagland TA, Wettemann RP (1984) Influence of elevated ambient temperatureafter breeding on plasma corticoids, estradiol and progesteronein gilts. Theriogenol 22(1): 15-24.
- Skarzynski DJ, Miyamoto Y, Okuda K (2000) Production of prostaglandinF (2alpha) by cultured bovine endometrial cells in response to tumornecrosis factor alpha: cell type specificity and intracellular mechanisms.Biol Reprod 62(5): 1116-1120.a
- Malayer JR, Hansen PJ, Gross TS, Thatcher WW (1990) Regulation ofheat shock-induced alterations in the release of prostaglandins by theuterine endometrium of cows. Theriogenol 34(2): 219-230.
- Badinga L, Collier RJ, Thatcher WW, Wilcox CJ (1985) Effects of Climaticand Management Factors on Conception Rate of Dairy Cattle in SubtropicalEnvironment. J Dairy Sci 68(1): 78-85.
- Wolfenson D, Thatcher WW, Badinga L, Savio JD, Meidan R, et al. (1995)Effect of heat stress on follicular development during the estrous cyclein lactating dairy cattle. Biol Reprod 52(5): 1106-1113.
- Ealy AD, Drost M, Hansen PJ (1993) Developmental changes in embryonicresistance to adverse effects of maternal heat stress in cows. J DairySci 76(10): 2899-2905.
- Wetteman RP, Bazer FW, Thatchar WW, Hoagland TA (1984) Environmentalinfluences on embryonic mortality. In: Xth International Conferenceon Animal Reproduction and Artificial Insemination, USA, p. 21-32.
- Ashburner M (1982) Heat Shock: from Bacteria to Man, Cold Spring HarborLaboratory, Cold Spring Harbor, New York, USA, p. 1-7.
- Danial NN, Korsmeyer SJ (2004) Cell Death: Critical Control Points. Cell116(2): 205-219.
- Lee AS (2001) The glucose-regulated proteins: stress induction and clinicalapplications. Trends in Biochem Sci 26(8): 504-510.
- Flint APF, Leat WMF, Sheldrick EL, Stewart HJ (1986) Stimulation ofphosphoinositide hydrolysis by oxytocin and the mechanism by whichoxytocin controls prostaglandin synthesis in the ovine endometrium.Biochem J 237(3): 797-805.
- Thatcher WW, Wolfenson D, Curl JS, Rico LE, Knickerbocker JJ, et al.(1984) Prostaglandin dynamics associated with development of the bovineconceptus. Anim Reprod Sci 7: 149-176.
- Putney DJ, Mullins S, Thatcher WW, Drost M, Gross TS (1989) Embryonicdevelopment in superovulated dairy cattle exposed to elevated ambienttemperatures between the onset of estrus and insemination. Anim ReprodSci 19: 37-51.
- Kobayashi Y, Wakamiya K, Kohka M, Yamamoto Y, Okuda K (2013) Summerheat stress affects prostaglandin synthesis in the bovine oviduct.Reprod 146(2): 103-110.